Úvod
Fosilní zdroje jsou hlavní suroviny pro energetické a organické sloučeniny, a jejich použití výsledky ve emisí skleníkových plynů spojených se změnou klimatu. Nadcházející klimatická havárie vyžaduje naléhavý přechod od fosilního hospodářství k bio-založené ekonomice, ve které se k výrobě paliv, chemikálií a materiálů používá spíše lignocelulózová biomasa než ropa. Dřevo je důležitým zdrojem lignocelulózové biomasy; skládá se hlavně ze sekundárně zahuštěných buněčných stěn bohatých na celulózu, hemicelulózy a lignin. Všechny tři polymery mohou být valorizovány v bio-založené ekonomice. Celulóza je zdrojem pro celulózu a papírenský průmysl a celulóza i hemicelulózy mohou být depolymerovány na jejich monosacharidy pro fermentaci na například bioethanol, kyselinu mléčnou a detergenty (Vanholme et al., 2013b). Jako lignin negativně ovlivňuje efektivitu zpracování dřeva k těmto aplikacím, stromy může být navržen tak, aby se hromadí méně ligninu, aby se stal přístupnější pro výrobu papíru a zkvasitelné cukry. Na druhé straně je lignin stále více považován za cennou složku v bio-založené ekonomice. Vskutku, vzhledem k tomu, že lignin je největší obnovitelných aromatické zdroj na Zemi, ekonomickou životaschopnost bio-rafinérie může být výrazně zvýšena, pokud lignin je také valorized, a používá se jako zdroj pro výrobu chemických látek (Holladay et al., 2007; Tuck et al., 2012; Davis a kol., 2013; Ragauskas et al., 2014; Li C. a kol., 2015; Van den Bosch a kol., 2015; Rinaldi a kol., 2016; Upton a Kasko, 2016; Schutyser a kol., 2018).
lignin polymer je složen z monolignols, které jsou produkovány phenylpropanoid a monolignol biosyntetické dráhy, sérií enzymatických reakcí počínaje deaminace fenylalaninu (Obrázek 1). Monolignoly jsou syntetizovány v cytoplazmě a translokovány do apoplastu, kde jsou dehydrogenovány na monolignolové radikály působením lakáz a peroxidáz (Berthet et al., 2011; Zhao a kol., 2013). Tyto monolignol radikály pak pár s sebou v různých kombinacích, vytváří řadu chemických vazeb jako aryl-éter dluhopisů (β-O-4), resinol dluhopisů (β-β), a phenylcoumaran dluhopisů (β-5) (Boerjan et al., 2003; Ralph et al., 2004; Vanholme et al., 2010). Nejčastější monolignols jsou hydroxycinnamyl alkoholy p-coumaryl, coniferyl, a sinapyl alkoholu, které generují H, G, a Y jednotek na jejich začlenění do lignin polymeru, respektive (Bonawitz a Chapple, 2010; Ralph et al., 2019; Vanholme a kol., 2019). Relativní příspěvek lignin stavební bloky se liší mezi taxony, vývojové fázi, tkáně a typu buněk, a dokonce i buněčná stěna vrstva; lignin z měkkého dřeva (nahosemenné) se skládá téměř výhradně z G jednotky s menší zlomek H jednotek, zatímco lignin z listnaté dřeviny (krytosemenné) se S jednotkami kromě G jednotky a stopy H jednotek (Boerjan et al., 2003; Vanholme et al., 2010, 2019). Kromě těchto tradičních monolignolů může být do ligninového polymeru začleněna řada dalších P-hydroxylovaných aromatických molekul na různé úrovně (Vanholme et al ., 2019).
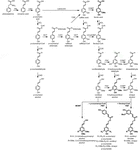
Figure 1. Lignin biosynthetic pathway. Alternative monomers and heterologously expressed enzymes are shown in bold. PAL, phenylalanine ammonia-lyase; C4H, cinnamate 4-hydroxylase; 4CL, 4-coumarate:CoA ligase; HCT, p-Hydroxycinnamoyl-CoA:quinate/shikimate-p-hydroxycinnamoyltransferase; C3’H, p-coumaroyl quinate/shikimate 3’-hydroxylase; CSE, caffeoyl shikimate esterase; CCoAOMT, caffeoyl-CoA O-methyltransferase; CCR, cinnamoyl-CoA reductase; F5H, ferulate 5-hydroxylase/CAld5H, coniferaldehyde 5-hydroxylase; COMT, kávová kyselina O-methyltransferáza; CAD, cinnamyl alkohol dehydrogenázy; FMT, feruloyl-CoA monolignol transferázy; PMT, p-coumaroyl-CoA monolignol transferázy.
Vzhledem k tomu, že lignin je hlavní vzpurnosti faktorem dřeva delignification procesy, velké výzkumné úsilí bylo věnováno odhalili lignin biosyntetickou dráhu, a na studie účinků poruchy v biosyntéze ligninu genů na lignin množství a složení, a na zpracování dřeva účinnosti. Zatímco modifikace v genech od těch kódujících transkripčních faktorů až po ty kódující oxidační enzymy vedly ke změně obsahu, složení nebo depozice ligninu (Eriksson et al., 2000; Li Y. H. a kol., 2003; Liang et al., 2008; Lu et al., 2013; Lin a kol., 2016; Xu et al., 2017; Yang a kol., 2017; Obudulu a kol., 2018), Tento přehled se zaměří na výsledky získané inženýrstvím biosyntetických genů ligninu.
technické Lignin Dráhy
Tabulka 1 poskytuje přehled různých studií na downregulated nebo mutované lignin biosyntetické geny v topol, borovice, eukalyptus a bříza, se výsledné efekty na zpracování dřeva efektivitu, když odhodlaný. Snížení aktivity jakéhokoli kroku biosyntetické dráhy ligninu, počínaje PAL až po CAD, může vést ke snížení obsahu ligninu (Tabulka 1). Několika parametry ovlivňují stupeň lignin snížení, jako cílový gen a stupeň down-regulaci aktivity enzymů, které zase závisí na účinnosti tlumení hluku postavit použité, velikost genové rodiny, a redundance v rámci genové rodiny. Obecně má downregulace kroků od C4H až po CCR za následek dramatičtější snížení množství ligninu (Hu et al ., 1999; Meyermans et al., 2000; Zhong et al., 2000; Li l. a kol., 2003; Jia a kol., 2004; Lu et al., 2004; Leplé et al., 2007; Coleman a kol.,2008a, b; Bjurhager a kol., 2010; Mansfield a kol., 2012; Ralph a kol., 2012; Min a kol., 2014; Van Acker a kol., 2014; Zhou a kol., 2015, 2018; Saleme et al., 2017; Xiang a kol., 2017) než downregulace F5H, COMT a CAD (Van Doorsselaere et al., 1995; Baucher a kol., 1996; Lapierre et al., 1999; Jouanin a kol., 2000; Van Acker a kol., 2014; Wang a kol., 2018). Redukce ligninu může být spojena se zvýšením S / G, například v C3 ‚ h- (Coleman et al., 2008a; Ralph a kol., 2012) a CCoAOMT-downregulated trees (Meyermans et al., 2000) nebo snížení poměru S / G, například v CSE – (Saleme et al., 2017) a COMT-downregulated trees (Van Doorsselaere et al., 1995; Lapierre a kol., 1999; Jouanin a kol., 2000). Je zajímavé, že bylo zjištěno, že topoly s nízkým ligninem 4CL-downregulated mají zvýšení S / G (Min et al ., 2014; Xiang a kol., 2017), pokles S / G (Voelker et al., 2010; Zhou a kol., 2015), nebo poměry srovnatelné s divokým typem (Hu et al., 1999; Li l. a kol., 2003). Tato odchylka nemůže být spojena s promotorem nebo metodou použitou pro downregulaci, což naznačuje, že rozdíly ve stupni umlčení, růstových podmínkách nebo vývojovém stavu ovlivňují tuto vlastnost. Na druhé straně, nejsilnější účinky na H/G/S lignin složení byly pozorovány pro stromy downregulated v C3 ‚ H a HCT, který vklad ligninu s velkými zvyšuje v H jednotky obsahu (Coleman et al., 2008a; Ralph a kol., 2012; Vanholme a kol., 2013a), zatímco stromy, které nadměrně exprimují F5H, produkují lignin silně obohacený o jednotky S (Franke et al ., 2000; Li l. a kol., 2003; Stewart a kol., 2009) a stromy, které jsou downregulovány v COMT, dramaticky snížily biosyntézu jednotek S (Van Doorsselaere et al., 1995; Lapierre a kol., 1999).

Tabulka 1. Přehled lesních stromů s modifikovanou expresí genů biosyntézy ligninu.
účinnost zpracování biomasy může být také změněna tím, že zvýšené inkorporaci molekul, které obecně představují menší komponenty v ligninu z wild-type rostliny. Začlenění ferulové kyseliny v CCR-nedostatečné stromy následek vznik acetalový dluhopisů v polymer lignin, které se snadno štěpí v kyselém biomasy pretreatments (Leplé et al., 2007; Ralph a kol., 2008; Van Acker a kol., 2014). Hladiny kyseliny ferulové v ligninu skutečně pozitivně korelovaly s vyšší účinností sacharifikace (Van Acker et al ., 2014). Začlenění 5-hydroxykoniferylalkoholu a 5-hydroxykoniferaldehydu do ligninu topolů s deficitem COMT (Van Doorsselaere et al ., 1995; Lapierre a kol., 1999; Jouanin a kol., 2000; Morreel et al., 2004; Lu et al., 2010) vede k vazbám benzodioxanu, které potenciálně brání kovalentním vazbám mezi ligninem a polysacharidovými hydroxylovými skupinami (Weng et al ., 2010; Vanholme a kol., 2012; Nishimura a kol., 2018). Na druhou stranu, COMT nedostatek také za následek více kondenzované lignin vzhledem k relativně vyšší úrovní kondenzované β-β a β-5 dluhopisy, a nižší hladiny β-O-4 dluhopisy, když S jednotkou frekvence kapek. Chemické rozvlákňování dřeva odvozené z topolů silně downregulated pro COMT za následek vyšší výnos buničiny vyvažuje zbytkový lignin obsah vlákniny. Tyto stromy měly nižší lignin a vyšší obsah celulózy (Jouanin et al., 2000). Na druhou stranu, topolů, které byly skromně downregulated pro COMT měl velký pokles výnos buničiny, pravděpodobně proto, lignin obsah zůstal normální, zatímco lignin měl vyšší frekvenci kondenzované dluhopisy, které negativně ovlivnily lignin extrakce (Lapierre et al., 1999; Pilát et al., 2002). Začlenění cinnamaldehydes v lignin polymeru v CAD-nedostatečné stromy výsledků v kratší lignin polymerních řetězců, tedy vyšší podíl volných fenolických koncových skupin, které zvyšují rozpustnost polymeru v alkalických. Začlenění cinnamaldehydes v lignin polymer pravděpodobně také snižuje kovalentní interakce alifatické řetězce s hemicelulózy, znovu vykreslování lignin více rozpustné. Kromě toho, vzhledem k rozšířené konjugovaný systém, který je generována, když cinnamaldehyd β-O-4 páry s další monomerní, aromatický ether dluhopisů zabudovaných cinnamaldehyd se stává více citlivé na alkalické štěpení (Lapierre et al., 1989; Van Acker a kol., 2017).
polymerace Ligninu je kombinatorické radikální spojovací proces, který umožňuje širokou škálu fenolických sloučenin být přirozeně začleněna do lignin polymer (Boerjan et al., 2003; Vanholme et al., 2019). Výzkumníci se pokusili přizpůsobit lignin množství a složení ke zlepšení zpracování biomasy podle exprese heterologní geny, jejichž cílem biosyntézy a začlenění různých kompatibilní fenolických sloučenin jako alternativní monolignols do lignin polymer (Ralph, 2006; Vanholme et al., 2012; Mottiar a kol., 2016; Mahon a Mansfield, 2018). Jedním z příkladů je zavedení genů kódujících enzymy, které jsou potřebné pro S jednotkou biosyntézy v borové; simultánní exprese F5H, COMT a CAD úspěšně představil S jednotkami v Pinus radiata (Wagner et al., 2015; Edmunds a kol., 2017). Zavedení genu kódující monolignol 4-O-methyltransferázy (MOMT4) do topolu vede ke vzniku 4-O-metylovaných coniferyl a sinapyl alkoholu, které nemohou být začleněny do rostoucího polymer lignin, protože jim chybí aromatické hydroxylové skupiny. To vede k zastavení polymerace ligninu a výsledky v stromy s nižším obsahem ligninu a vyšší zcukernatění účinnost (Bhuiya a Liu, 2010; Cai et al., 2016). Topoly byly také navrženy tak, aby obsahovaly esterové vazby v páteři ligninového polymeru. Estery coniferylferulátu byly zavedeny do polymeru expresí genu FERULOYL-COA: MONOLIGNOL transferázy (FMT) odvozeného od Angelica sinensis (Wilkerson et al ., 2014), což vede ke zlepšení účinnosti sacharifikace za různých podmínek předúpravy (Wilkerson et al ., 2014; Kim a kol., 2017; Bhalla a kol., 2018) a zlepšená účinnost rozvlákňování kraft ve srovnání s divokým typem (Zhou et al ., 2017). Monolignol p-coumarate estery byly také vyvinuty v topol, přes vyjádření rýže, p-COUMAROYL-CoA:MONOLIGNOL TRANSFERÁZY (PMT) genu, což má za následek vyšší frekvenci rezistentních interunit dluhopisů a vyšší frekvence G a S terminální jednotky s bezplatným fenolické skupiny (Smith et al., 2015; Sibout a kol., 2016). Zatímco u Arabidopsis vedla heterologní exprese PMT ke sníženému množství ligninu doprovázenému zvýšenou účinností sacharifikace (Sibout et al., 2016), nedošlo k žádnému snížení množství ligninu v topolu a účinnost sacharifikace nebyla stanovena (Smith et al., 2015).
Zatímco několik modifikací ligninu množství a složení bylo prokázáno, že poskytují vylepšení ve zpracování biomasy, tyto změny jsou často doprovázeny biomasy výnos trest (Leplé et al., 2007; Wadenbäck et al., 2008; Wagner a kol., 2009; Voelker et al., 2010; Stout et al., 2014; Van Acker a kol., 2014; Sykes et al., 2015; Zhou a kol., 2018). Nedávná metastuda narušila 21 genů biosyntézy ligninu v P. trichocarpa a komplexně integroval výsledky transkriptomických, proteomických, fluxomických a fenomických dat 221 řádků. Autoři dospěli k závěru, že růst stromů není spojen s množstvím ligninu, složením podjednotky nebo specifickými vazbami (Wang et al., 2018), ale spíše koreloval s přítomností zhroucených xylemových cév (Coleman et al., 2008a, b; Wagner a kol., 2009; Voelker et al., 2010; Vargas a kol., 2016; De Meester et al., 2018), aktivace cesty integrity buněčné stěny (Bonawitz et al ., 2014) a / nebo akumulace chemických inhibitorů (Gallego-Giraldo et al., 2011; Muro-Villanueva et al., 2019).
Vzhledem k tomu, že značné úsilí bylo vynaloženo na snížení obsahu ligninu downregulation ligninu biosyntetické geny, studium na upregulace lignin dráhy a nadprodukce ligninu byly vzácné. Zprávy o nadměrné expresi F5H skutečně ukazují nezměněný nebo dokonce pokles obsahu ligninu (Huntley et al ., 2003; Li l. a kol., 2003; Stewart a kol., 2009; Mansfield a kol., 2012; Edmunds a kol., 2017). Nadměrná exprese CAD a COMT vedla spíše k umlčení genů než k upregulaci nebo nebyl zjištěn žádný účinek na hladiny exprese (Baucher et al., 1996; Lapierre et al., 1999; Jouanin a kol., 2000; Leplé et al., 2007; Van Acker a kol., 2014). Nadměrné expresi z R2R3-MYB transkripční faktory PtoMYB92, PtoMYB216, a PtoMYB74 vše vyústilo v další xylem vrstev, tlustší buněčné stěny xylému, stejně jako mimoděložní depozice ligninu, a rostliny nahromaděné 13-50% více ligninu (Tian Q. et al., 2013; Li C. F. et al., 2015; Li a kol., 2018). Zvýšená exprese MYB linky constitutively upregulován na biosyntéze ligninu cestě genů, a zatímco rostliny se zvýšenou expresí MYB92 a MYB74 měl biomasy trest, zvýšená exprese MYB216 za následek rostliny s až o 50% více ligninu a ne vývojové fenotyp. Jako lignin je stále více považována za důležitý zdroj pro udržitelné výroby chemických látek (Cao et al., 2018) by mělo být dále prozkoumáno inženýrství rostlin nadprodukovajících lignin.
Polní studie
výše uvedené příklady jasně ukazují, že inženýrství ligninu prostřednictvím down-nebo upregulace genů fenylpropanoidní dráhy – nebo exprese heterologních genů-má potenciál zvýšit účinnost zpracování lignocelulózové biomasy. Z praktických a regulačních důvodů většina studií uvádí údaje získané z analýzy stromů pěstovaných ve skleníku. Experimenty se stromy pěstovanými ve skleníku však obvykle nezohledňují vývojové procesy, jako je zastavení růstu a dormance. Kromě toho skleníkové experimenty neposkytují dostatečný vhled do interakce upravené rostliny s faktory prostředí, jako je typ půdy, vítr a patogeny. Pochopení těchto interakcí je důležitým krokem při překladu výsledků výzkumu směrem ke komerčním aplikacím. Vskutku, soubor prací vytvořených studiemi, pro které bylo uděleno povolení k zavedení polních zkoušek, zdůrazňuje důležité rozdíly ve fenotypu mezi skleníkem – a stromy pěstované v terénu. Tabulka 1 shrnuje zprávy o polních pokusech provedených se stromy 4CL, CCoAOMT, CCR, COMT a CAD downregulated.
potvrzení potenciálu modifikované lignocelulózové biomasy jako substrátu pro aplikace, několik ligninem upravených polních pokusů pěstovaných stromů ukázalo zlepšení zpracování dřeva. Topoly downregulated pro CCoAOMT pěstovány po dobu 5 let v terénní studii v Pekingu (Čína), ukázaly zvýšené glukózy a xylózy uvolnění po zcukernatění (Wang et al., 2012). Topoly downregulated pro CCR a pěstovány v oblasti hodnocení ve Francii, se ukázala být přístupnější chemické kraft rozvlákňování (Leplé et al., 2007). Další dvě pole studiích provedených ve Francii a Belgii s CCR-downregulated topoly za následek až 160% zlepšení v produkci etanolu v současné zcukernatění a fermentace (SSF) testy; nicméně rostliny měly až o 50% snížení biomasy (Van Acker et al., 2014). Nadějné výsledky vykázaly i terénní zkoušky s CAD-downregulovaným topolem. Tyto stromy vykazovaly o něco méně ligninu než divoký typ a ukázaly se lépe přístupné delignifikaci kraft (Lapierre et al., 1999). Důsledně, stejné linky pěstuje ve větším měřítku polní pokusy ve Francii a Spojeném Království ukázaly mírný pokles v množství ligninu a zlepšení kraft rozvlákňování považováno za komerčně důležité, protože rostliny potřebné 6% méně alkalické dosáhnout delignification podobná wild-type stromy (Pilát et al., 2002).
nicméně protichůdné zprávy o výtěžku biomasy a účinnosti následného zpracování naznačují, že tyto parametry jsou vysoce ovlivněny faktory prostředí. Zatímco terénní studie provedená v Číně s použitím 4CL downregulovaných topolů zjistila, že i při 28% poklesu obsahu ligninu ve srovnání s divokým typem měly stromy asi 8% zvýšenou výšku (Tian X.M. et al., 2013), v souladu se skleníkovými studiemi (Hu et al., 1999), jiné polní pokusy zjistily, že topoly 4CL-downregulated snížily biomasu a někdy byly dokonce zakrslé (Voelker et al., 2010; Stout et al., 2014; Marchin et al., 2017). Zprávy se také rozcházejí, pokud jde o efektivitu zpracování dřeva získaného z těchto topolů pěstovaných v terénu 4CL-downregulated. Zatímco až 100% nárůst cukru zotavení, bylo zjištěno na 4CL1-downregulated stromy (35S-řízený antisense 4CL construct) pěstované v horské lokalitě ve Spojených Státech (Xiang et al., 2017), data získaná z terénních studií provedených v Oregonu (usa) zjistili, že Pt4CL1 promotér-řízený „antisense“ umlčel 4CL1 topoly neměl žádné zlepšení v zcukernatění účinnost ve srovnání s divokým typem (patří obchodní centrum, přístup et al., 2010). Navíc, dlouhodobé studie v Wenling (Čína), zjistil, že 4CL-downregulated topoly neprokázalo významné zlepšení v cukru výnos ve srovnání s divokým typem (Wang et al., 2012). V obou posledně uvedených případech, stromy vykázaly mírné snížení množství ligninu, které se nepromítly do vyšší efektivitu zpracování, potenciálně, protože vyšší koncentrace extrahovatelných látek, které by mohly interferovat s enzymatickou aktivitu (patří obchodní centrum, přístup et al., 2010).
Field trial studie prokázaly, že faktory životního prostředí mohou ovlivnit dřevnatění a obnovit rysy wild-type úrovně ve srovnání s úrovní dosáhnout při stejné rostliny byly pěstovány ve skleníku. Zatímco 4CL-downregulated stromy snížil obsahem ligninu, když pěstovány ve skleníkových podmínkách, analýza stejný 4CL antisense topoly, ale pěstované na poli, se často ukázalo, že obsah ligninu byla zvýšena ve srovnání s skleníku pěstované stromy a někdy dokonce obnovena wild-type úrovni (Stout et al., 2014; Xiang a kol., 2017). Podobně byly hladiny ligninu mnohem méně sníženy u topolů s deficitem CCR, když byly pěstovány na poli, ve srovnání s tím, kdy byly pěstovány ve skleníku (Van Acker et al ., 2014). Alespoň u topolů s nedostatkem CCR je možné, že vyšší úroveň lignifikace polních stromů je způsobena skutečností, že vzorky dřeva byly odebrány během zimy. Když strom přestane růst na podzim, stromy stále mít čas, aby plně lignify jejich buněčné stěny v době, strom vstoupí do dormance, ve srovnání s skleníku-vzrostlé stromy, které vyvíjejí nové xylem nepřetržitě. Ukázalo se také, že složení ligninu se liší mezi stromy s nízkým ligninem pěstovanými ve skleníku a na poli. 4CL-downregulated topoly pěstované na poli v Severní Karolíně měly lignin s nižším S / G, než když byly stejné linie pěstovány ve skleníku (Stout et al ., 2014).
celkově tyto výsledky ukazují, že údaje získané ze skleníkových stromů nelze snadno extrapolovat na stromy pěstované v terénu, což podporuje potřebu pokusů v terénu na různých místech. Některé linky představila výnosu penále činí je méně zajímavé pro aplikace, které zdůrazňují potřebu lepší pochopení molekulární základ, z výnosu penále a rozvoj strategií k překonání tohoto problému.
bylo prokázáno, že Lignin hraje důležitou roli v rezistenci na patogeny (Miedes et al., 2014; Zhao a Dixon, 2014), a hraje klíčovou roli při umožnění závodu přepravovat vodu. To naznačuje, že modifikace ligninu by mohly mít vliv na toleranci stresu rostlin. Zatímco další vyšetřování je potřebné, aby plně adresa tato možnost, down-regulaci 4CL, COMT, a CAD v poplar ne dramaticky změnit krmení výkon list-krmení býložravců (Tiimonen et al., 2005; Brodeur-Campbell et al., 2006; Hjalten et al., 2013). Účinek downregulation COMT a CAD v topol na rostliny-hmyz interakce byla také hodnocena na poli vzrostlé stromy, a bylo prokázáno, že lignin-modifikované stromy měl normální výskyt návštěvě a krmení hmyzu, stejně jako normální odpovědi na mikrobiální patogeny (Pilát et al., 2002; Halpin et al., 2007). Tyto výsledky naznačují, že stromy s modifikovaným ligninem nemusí nutně trpět více než rostliny divokého typu škůdci a chorobami. Nicméně, profilování endosphere bakteriální mikrobiomem dřevo sklizené z pole-pěstuje, CCR-downregulated topoly demonstrovány posuny v bakteriální společenství, pravděpodobně z důvodu změněné množství jednotlivých fenolických metabolitů v xylému (Beckerse et al., 2017).
vzhledem k roli ligninu ve funkci a struktuře xylemu byly hodnoceny vodní vztahy několika topolů modifikovaných nízkým ligninem. 4CL-downregulated topoly bylo zjištěno, že mají sníženou hydraulickou vodivost, potenciálně interferující s růstem rostlin (Marchin et al ., 2017). Hydraulický stres experimenty s topoly downregulated pro CCR, COMT nebo CAD ukázala, že tyto rostliny mají nižší odolnost proti kavitaci, při zachování normální xylem hydraulické vodivosti a vodní dopravy (Awad et al., 2012). Tyto výsledky naznačují, že růst nízko ligninových mutantů může být ovlivněn dostupností vody. Stejně jako u každého nového hybridu získaného z klasického šlechtění jsou nutné terénní testy k vyhodnocení výkonu v terénu a tolerance stresu stromů upravených ligninem.
vyhlídky na inženýrství ligninu v lesních stromech
zdá se, že výkonnost rostlin z ligninu je silně ovlivněna podmínkami prostředí. Je však nejasné, zda pozorované rozdíly mezi emisí skleníkových pěstované a poli vzrostlé stromy, nebo mezi stromy pěstované v různých terénu, jsou výsledkem různých úrovních genové potlačení nebo z interakce inženýrství vlastnost prostředí (GxE). Nestabilní downregulace je ve skutečnosti nedostatkem technik umlčování genů, které jsou založeny na RNAi. O tom svědčí pozorování variace v červené xylem fenotyp, který je pozorován při konkrétní biosyntézy ligninu genů, jako jsou CAD, COMT, nebo CCR, jsou downregulated. Červené zbarvení xylem často není jednotné v celém xylemu, ale spíše se objevuje v náplastech, které odrážejí variabilní úrovně umlčení genů (Leplé et al., 2007; Voelker et al., 2010; Van Acker a kol., 2014; Obrázek 2). Kromě toho, použití metod umlčování genů může potenciálně vést k souběžnému umlčení blízce příbuzných členů genové rodiny-možná v různé míře-zakalení interpretací a maskování účinků downregulace jednotlivých genů.

Obrázek 2. Nerovnoměrná regulace genu RNAi. Nerovnoměrný červený fenotyp xylem pozorovaný na kmenech topolů s deficitem CCR (vpravo) pěstovaných v polním pokusu v Belgii. Červený xylem označuje oblasti DOWNREGULACE CCR. Dřevo ze stromů divokého typu je bělavé (vlevo).
Tyto problémy mohou být nyní snadno překonat pomocí CRISPR-based editace genové technologie, které umožňují stabilní loss-of-function mutace (knock-out) v konkrétních cílových genů, což pitva funkce jednotlivých genů v rodinách. Například cílení jednotlivých 4CL gen členy rodiny v topol ukázal, že 4CL1 souvisí s dřevnatění, vzhledem k tomu, že 4CL2 je zapojen v proanthocyanidin výroby (Zhou et al., 2015). Kromě knock-out alel umožňuje editace genů na bázi CRISPR také vytvářet nové alely, které poskytují částečné snížení enzymové aktivity. To otevírá možnost doladit úroveň zbytkové enzymové aktivity a procesy, výnos trest, který je často pozorován, když lignin částka klesne pod prahovou úroveň. Další slibnou cestou pro lignin inženýrství v lese stromy možné pomocí CRISPR-na základě genomu inženýrství je simultánní editace více geny (alely stohování) pro optimalizaci zpracování biomasy účinnosti, např. v Arabidopsis, kde stohování transaldoláza (tra) a comt mutace, c4h a comt mutace, nebo 4cl a comt mutace za následek aditivní a synergické zlepšení v zcukernatění účinnost (de Vries et al., 2018). Vskutku, systémový přístup v P. trichocarpa předpovídá, že současné downregulation PAL a CCoAOMT, nebo PAL, C3 ‚ H a CCOAOMT podstatně zlepší vlastnosti dřeva a cukru vydání (Wang et al., 2018).
použití CRISPR-based editace genomu v strom zlepšení pro výrobu buničiny a papíru a bio-rafinerie průmysl, stejně jako pro výrobu platforma aromátů z hydrogenolytic odbourávání ligninu, bude velmi cenné, když tato technologie je strategicky v kombinaci s jinými technikami pro šlechtění (Obrázek 3). Velké rozdíly v množství ligninu a složení S/G již existují v přirozených populacích lesních stromů (Studer et al ., 2011). Vzhledem k tomu, že obě vlastnosti ovlivňují uvolňování glukózy při sacharifikaci (Yoo et al ., 2018), využívající tuto genetickou rozmanitost konvenčním šlechtěním, za pomoci genomových asociačních studií (GWAS) (Porth et al., 2013; Fahrenkrog a kol., 2017; Liu a kol., 2018), chov se vzácnými vadnými alelami (BRDA) (Vanholme et al., 2013a) nebo genomický výběr (Yin et al., 2010; Muchero a kol., 2015; Pawar et al., 2018; Xie a kol., 2018), je cennou strategií pro získání linek, které zlepšily efektivitu zpracování dřeva. Kdysi elitní stromy jsou získané pomocí těchto metod šlechtění, genetické inženýrství a CRISPR-based editace gene specifických genů je velmi slibnou cestou k dalšímu zlepšení těchto elitních genotypů bez rozbití jejich genetické konstituci a bez zdlouhavé chovných cyklů. Vzhledem k hrozící klimatické havárii nemáme více času ztratit při přijímání těchto nových šlechtitelských technik v našem závodě s biobazovanou ekonomikou.
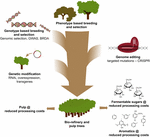
obrázek 3. Genetické zlepšení lesních stromů kombinací šlechtitelských nástrojů. Pro urychlení genetického zlepšení lesních stromů pro aplikace buničiny a biorefinery je třeba elegantně kombinovat klasické a nové šlechtitelské nástroje. Klasický šlechtění zahrnuje fenotypový výběr stromů pro řízené kříže, následovaný fenotypovým výběrem. S příchodem sekvence genomu, informace z mnoha lesních stromů, nové strategie, jako jsou Genomické Selekce, Genome Wide Association Studies (GWAS) a Chov Vzácných Vadné Alely (BRDA) byly vyvinuty k urychlení zachytit a obohacení DNA polymorfismy spojené s prospěšné vlastnosti. Úpravy genomu založené na CRISPR umožňují modifikovat genom způsobem, který napodobuje přirozené polymorfismy. Genetická modifikace zahrnuje stabilní integrace cizí DNA do stromu k nadprodukci (e) enzym(y), nebo downregulate (a) gen(y). Kombinace klasických a nových šlechtitelských technik je nutná k zajištění dostatečného množství vysoce kvalitního dřeva pro společnost.
Autor Příspěvky
Všichni autoři uvedených mít podstatné, přímé a intelektuální příspěvek k práci, a to schválil ke zveřejnění.
financování
uznáváme částečné financování z projektu IWT-SBO BIOLEUM (Grant č. 130039) a SBO-FISH prostřednictvím projektu ARBOREF. AC získala finanční prostředky z FWO a Programu Evropské unie pro výzkum a inovace Horizon 2020 v rámci grantové dohody Marie Skłodowska-Curie č. 665501. LdV byl financován Institutem pro podporu inovací prostřednictvím vědy a techniky ve Flandrech (IWT-Vlaanderen) pro predoctoral fellowship.
Prohlášení o střetu zájmů
autoři prohlašují, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly být vykládány jako potenciální střet zájmů.
poděkování
Děkujeme Annicku Bleysovi za přípravu tohoto rukopisu k odeslání.
Boerjan, W., Ralph, J., and Baucher, m. (2003). Biosyntéza ligninu. Annu. Rev Plant Biol. 54, 519–546.
PubMed Abstraktní | Google Scholar
Lapierre, C., Pollet, B., Petit-Conil, M., Toval, G., Romero, J., Pilát, G., et al. (1999). Strukturální změny z ligninů v transgenních topoly s depresivní cinnamyl alkohol dehydrogenázy nebo kyselina kávová kyselina O-methyltransferáza aktivity mají opačný vliv na efektivitu průmyslové kraft rozvlákňování. Rostlinný Fyziol. 119, 153–163.
PubMed Abstract / Google Scholar
Ralph, J. (2006). „Co dělá dobrou náhradu monolignolu?, „ve vědě a tradici rostlinné buněčné stěny-biosyntéza – struktura a funkce, ed. T. Hayashi (Boca Raton, FL: BrownWalker Press).
Google Scholar