Úvod
Úspěšné interakce s naší smyslové prostředí vyžaduje složitou rovnováhu dva pozornostní mechanismy výběru—to top-down a bottom-up. Mířím k uličce v místním supermarketu s cílem vyzvednout několik potřebné ingredience pro mango salát, zapojuje nasazení dobrovolných cíl-režie, nebo top-down, pozornostní systém tak, že budete aktivně vyhledávat všechny potřebné ingredience mezi množstvím vyrábět volby. Pokud však uslyšíte vyzvánění mobilního telefonu, s největší pravděpodobností upoutá vaši pozornost a přeruší vyhledávání. Takové přerušení dojde v bottom-up, nebo stimulační-řízený, móda, kdy pouhý význam stimulu, skutečnost, že prsten se liší od ostatních zvuků v prostředí, považuje za hodné výběru. Popsaný scénář podtrhuje význam cílené a stimul-řízený výběr pro chování, a poukazuje na jemnou rovnováhu, která musí existovat mezi dva pozornostní systémy, aby se zabránilo „tunelové vidění“ na jedné straně a naprostou neschopnost se soustředit na další.
výběr shora dolů a zdola nahoru: Chování
Několik desetiletí behaviorální výzkum byl věnován prokazující, že rozložení pozornosti může být řízen záměry pozorovatele, stejně jako o důležitosti fyzického stimulu. Hodně behaviorálních důkazů pro alokaci pozornosti shora dolů a zdola nahoru bylo rozsáhle přezkoumáno jinde (Johnston a Dark, 1986; Egeth a Yantis, 1997). Abychom to shrnuli, studie prokazující účinky kontroly pozornosti shora dolů ukazují, že pozornost může být úspěšně přidělena prostorovým místům, funkcím,objektům atd., po přítomnosti exogenních nebo endogenních podnětů (Eriksen a Hoffman, 1972; Posner, 1980; Posner a kol., 1980), nebo očekávání nastavená buď předchozí znalosti nebo události stimulu (Shaw, 1978; Moore a Egeth, 1998; Geng a Behrmann, 2002, 2005; Shomstein a Yantis, 2004a; Drummond a Shomstein, 2010). Důkazy na podporu zdola nahoru pozornostní rozdělení se spoléhal na různé pozornostní zachytit paradigmata, v nichž účastníci jsou zapojeni v top-down hledat a jejich pozornost je obrácena k úkolu-irelevantní podněty, které prokazují, že pozornost je zajat funkce u jednoho dítěte (jedinečné položky; Yantis a Jonides, 1990; Theeuwes, 1991; Folk et al., 2002) a náhlý nástup (Yantis a Jonides, 1984; Theeuwes, 1991; Koshino et al., 1992; Juola a kol., 1995).
Vzhledem k tomu, že většina raných studií se soustředilo na prokázání důkazy pro top-down a bottom-up pozornostní výběr, nejnovější studie přesunula svou pozornost na zkoumání, jak dva pozornostní výběr systémů komunikovat. Tato linie vyšetřování je poháněný pozorování, že v zájmu efektivně vyberte úkol-relevantní informace (např., ingredience pro salát) musí aktivně inhibují úkol-irelevantní informace, které by jinak odvrátit pozornost od úkolu. Odvrácenou stranou této logiky je, že čím méně je zaměřeno na informace týkající se úkolů, tím více bude následovat zachycení. Bylo prokázáno experimentálně, že pozornost státu pozorovatele předpovídá, jaký typ informací, a do jaké míry, bude nakonec zachytit pozornost (Folk et al., 1992, 2002; Bacon and Egeth, 1994; Gibson and Kelsey, 1998). Například Folk et al. (2002) ukázal, že při hledání červeného písmene bude pozorovatel snadněji zachycen irelevantním podnětem na periferii, pokud je tento podnět červený nebo nějakým způsobem odpovídá cílové šabloně. Protože pozorovatel je top-down control nastavení jsou nastaveny na hledání červené funkce, stimulační, která je červená, je pravděpodobné, že upoutat pozornost a potenciálně zasahovat do řízení shora-dolů. Tak, s zachytit úkol, pozornostní strategie vyhledávání lze odlišit od sebe různé podobnosti úrovní mezi stimulační vlastnosti cíle a distraktorů. Čím více je cíl podobný distraktoru, tím obtížnější je, aby se pozorovatel vyhnul zachycení.
Role parietálního laloku při výběru shora dolů a zdola nahoru: Neurologický
Různé neuroimaging technik, za předpokladu, silný důkaz pro zapojení parietální kůry v top-down a bottom-up orientace, s důkazy přezkoumány značně jinde (Corbetta a Shulman, 2002, 2011; Behrmann et al., 2004). Bylo prokázáno, že oblasti, nejčastěji aktivován následující top-down podněty k účasti na konkrétní místa, funkce nebo objekty, které jsou umístěny podél hřbetní části temenní kůry. Takové oblasti patří nižší parietální lalůček (IPL), dorsomedial regiony označovány jako vynikající parietální lalůček (SPL), stejně jako další mediální oblasti podél precuneus gyrus (Yantis et al., 2002; Giesbrecht et al., 2003; Liu a kol., 2003; Yantis a Serences, 2003; Obrázek 1). Několik úkolů shora dolů bylo ukázáno, aby se úspěšně zapojily dorzální oblasti parietální kůry, konkrétně ty, které zahrnují prostorové (Kastner et al ., 1999; Corbetta a kol., 2000; Hopfinger a kol., 2000; Shomstein a Behrmann, 2006; Chiu a Yantis, 2009; Greenberg a kol., 2010), jakož i neprostorové posuny pozornosti (Giesbrecht et al., 2003; Yantis a Serences, 2003; Shomstein a Yantis, 2004b, 2006; Tamber-Rosenau et al., 2011).
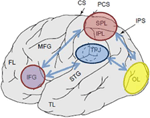
Obrázek 1. Schematické zobrazení příslušných anatomických orientačních bodů promítaných na boční povrch lidského mozku. Vynikající parietální lalůček (SPL) a dolní temenní lalůček (IPL) jsou regiony, ve hřbetní části temenní kůry podřídilo top-down pozornostní orientace. Temporo-temenní junkce (TPJ) je oblast ve spodní temenní kůře podřídilo zdola nahoru, pozornost, orientace. Oba, SPL a TPJ, jsou myšlenka k vyvolání řídicí signály zodpovědný za následné pozornostní modulací pozorován přes smyslové regionů, v tomto případě modulační (označeny tmavě modré šipky) vizuálně evokované aktivity v týlním laloku (OL). Navíc, oblasti podél dolní čelní závit (IFG) a dolní čelní křižovatce (IFJ) jsou myšlenka sloužit jako konvergence oblastí pro stimul-řízený a top-down pozornostní kontroly (označené světle modré bi-směrové šipky).
V typické úlohy zaměřené, aby se zapojily top-down pozornostní rozdělení, jednotlivci jsou zobrazeny dva rapid serial visual prezentace (RSVP) proudy umístěn periferně a jsou nejprve poučeni, aby sledovat jeden proud na signál (např. číslice mezi proud dopisů). Identita tága označuje, zda subjekt musí udržovat pozornost na aktuálním proudu nebo přesunout pozornost na druhý proud (Yantis et al., 2002; Yantis and Serences, 2003). V takových paradigmatech jsou pozorovány dva hlavní nálezy. První souvisí se zvýšenou aktivací v rámci smyslové regionů, zastupující na-the-moment zúčastnili umístění (např, zvýšená aktivita v levé primární zrakové regionů, kdy právo RSVP proud se zúčastnil). Toto zjištění poskytuje přesvědčivé důkazy, že účastníci se účastní na konkrétní místo a že pozornost moduluje sílu smyslové reakce (viz Obrázek 1; Moran a Desimone, 1985; O’Craven et al., 1997). Druhé zjištění souvisí s pozorováním, že hřbetní oblasti parietálního laloku jsou selektivně aktivovány posuny pozornosti shora dolů. Je zjištěno, že SPL/IPL timecourse činnosti je přechodné povahy, což naznačuje, že tato oblast parietální kůra je zdrojem stručný attentional control signál k posunu pozorný státy v top-down způsobem (Yantis et al., 2002).
Několik fMRI studií dokumentují, že bottom-up pozornostní zachytit, zprostředkované stimulační význam a/nebo význam, je subserved tím, temporo-parietálního křižovatky (TPJ; Obrázek 1). Například, když subjekty navštěvují a monitorují změnu vizuálního nebo sluchového podnětu, prezentovaného současně, je zvýšena aktivace oblastí TPJ parietálního laloku. Kromě zjevné citlivost na důležité podněty, TPJ je také aktivován v reakci na potenciálně román (neočekávané nebo málo časté) události, kdy je organismus zapojen do neutrální chování kontextu, nebo když se zabýval úkol (Marois et al., 2000; Downar et al., 2002; Serences et al., 2005; Corbetta a kol., 2008; Asplund et al., 2010; Diquattro a Geng, 2011; Geng a Mangun, 2011). Tato aktivace probíhá nezávisle na modalitu (sluchové, hmatové a vizuální), ve kterém je vstup nezaujme, což odráží multisenzorický povahu TPJ (ale viz Downar et al., 2001).
typický úkol zkoumá neurální mechanismus bottom-up pozornostní zachytit, účastníci jsou prezentovány s RSVP proud položky ve středu displeje, a jsou požádáni, aby určili, předem definovaný cíl (např. identifikovat červené písmeno prezentovány v rámci RSVP proud white non-cíle). Některé část zkoušky obsahuje úkol-nepodstatné charakteristické distraktoru prezentovány v různých časových intervalech před nástupem cíl, zatímco jiné testy obsahují pouze hlavní distraktoru (tj. bez cíle). „Cíl-distraktoru“ studiích jsou používány za účelem stanovení rozsahu zachytit, což ukazuje, že úkolem-irelevantní distraktoru je ve skutečnosti charakteristické tím přinese snížení cílové přesnost. „Distraktoru-v-izolaci“ zkoušky jsou použity pro další analýzy, protože tyto zkoušky umožňují vyšetření aktivita vyvolala na výběžku distraktoru bez kontaminace od cíle související procesy. Z takových paradigmat vyplývá několik důležitých zjištění. Za prvé, když jsou distraktory prostorově odděleny od cílového umístění, zachycovací distraktory jsou doprovázeny zvýšenou kortikální aktivitou v odpovídajících oblastech smyslové kůry (např. Tyto výsledky poskytují silný důkaz, že během zachycení je prostorová pozornost ve skutečnosti zachycena na prostorové místo obsazené distraktorem (Serences et al ., 2005). Za druhé, ventrální oblasti parietální kůry, zejména v TPJ, jsou selektivně aktivovány nedobrovolnými posuny pozornosti zdola nahoru. Jen jako činnost v rámci SPL pro top-down orientace, timecourse činnosti pozorovány v průběhu TPJ je přechodné povahy, což naznačuje, že tato oblast je zdrojem stručný attentional control signál k posunu pozornosti ve zdola nahoru.
Je třeba poznamenat, že zatímco tato recenze je zaměřena na řešení kognitivních funkcí, zadní parietální kůra, ostatní regiony, zejména ty, ve frontální mozkové kůry jsou také přijati pro top-down a bottom-up pozornostní rozdělení. Tyto regiony patří ventrální frontální kůře (VFC), frontální oční pole (FEF), nižší čelní křižovatce (IFJ), a dolní čelní závit (IFG; Corbetta a Shulman, 2002, 2011; Serences et al., 2005; Asplund et al., 2010; Diquattro a Geng, 2011).
Role parietálního laloku při výběru shora dolů a zdola nahoru: Neuropsychologie
Historicky vědci spoléhali kriticky na neuropsychologické studie pacientů s hemispatial zanedbávání (porucha prostorové rozdělení pozornosti na levé hemi-space) získat vhled do kognitivní funkce spojené s parietální lalok. V klasické neuropsychologické literatuře byla parietální kůra obecně považována za místo primární léze pro hemispatiální zanedbávání. Tento pohled, podrobně zpracovaný časnými vědci (Critchley, 1953; McFire a Zangwill, 1960; Piercy, 1964) jasně rozpoznal souvislost mezi parietální lézí a následným zanedbáváním. Tento pohled byl do značné míry držel přes 1980, kdy Posner a kolegové (1984) používá skryté visuospatial přehrávání paradigma ukazují, že poškození mozku vytváří deficit v „uvolnit“ provoz (zasouvání pozornost z jednoho místa a přesouvá do jiného), když cíl je kontralaterální k lézi. Nicméně, i přes tento významný pokrok v porozumění nervový základ pozornost a konkrétně „uvolnit“ roli parietální kůře, jejich zjištění, že jeden kortikální stránky (parietální kortex) a jeden funkční schopnosti („odpojit“). V kontrastu s tímto více monolitických přístup k mozku (parietální kortex) a chování (pozornost, uvolnění), nedávné chování a neurologický práce (přezkoumána výše a jinde) naznačuje, že oba kortikální oblasti a související pozornostní chování lze rozčlenit do kvalitativně různých profilů.
Vzhledem k segregaci kortikální sítě do top-down a bottom-up procesy, zřejmý předpověď je, že poškození superior části parietální lalůček (subsuming SPL) by měl přinést deficit ve cílená pozornost, orientace, vzhledem k tomu, že poškození dolní části parietální lalůček (subsuming TPJ) by mělo za následek deficit spojených s stimulus-driven pozornost zachytit. Do té míry, že tyto korespondence chování mozku byly zkoumány v neuropsychologické literatuře, tato předpověď není zjevně potvrzena. Například, klinické příznaky hemispatial zanedbávání jsou silně spojené s poškozením nižší části parietální lalok, který zahrnuje TPJ, spíše než vynikající porce jako SPL (Friedrich et al., 1998; Shomstein a kol., 2010; Corbetta a Shulman, 2011). To je poněkud v rozporu s neuroimaging literatury, což naznačuje, že role TPJ je v zachycení pozornosti, spíše než v dobrovolné orientace pozornosti, domény, ve kterém zanedbávání pacientů se zdají mít největší potíže. Pro další zkomplikování bylo zjištěno, že léze, které zahrnují výhradně SPL, jen zřídka produkují klinické důkazy o zanedbání (Vallar a Perani, 1986). Další nedávná studie s pacienty s lézemi zaměřenými primárně na TPJ a STG, ale zachovanými SPL, Corbetta et al. (2005) ukázal, že prostorové zanedbávání, stejně jako jeho využití, byla spojena s obnovení činnosti v obou ventrální temporo-parietální a dorzální parietální regiony (viz Corbetta a Shulman, 2011 na recenzi). Zatímco zajímavé a vzrušující ve svých závěrech, tato poslední studie nerozlišuje relativní příspěvek dorzální a ventrální dráhy pro různé typy pozornost, protože pacienti byli testovány pouze na variantu Posner skryté prostorové pozornost cuing úkol, úkol, který je myšlenka, aby se zapojily oba top-down a bottom-up, pozornost, orientace.
rozlišovat mezi cílem-řízený attentional control a charakteristické pozornostní zachytit a zkoumat jejich mapování na SPL a TPJ, respektive nedávné studie přijala dvě paradigmata behaviorální, každý po jedné z těchto forem pozornost (Shomstein et al., 2010). Zkoumat integritu top-down pozornost, orientace v pacientů, top-down úkol byl použit vyžadují, aby účastníci směny prostorové pozornosti mezi prostorově odděleny RSVP proudy (úkol, který byl úspěšně použit k prokázání SPL aktivace na fMRI studií (Yantis et al., 2002)). Podobně, za účelem zkoumání schopností orientace pozornosti zdola nahoru pacientů, varianta Folk et al. (2002) podmíněné zachytit paradigma byl zaměstnán ve které účastníci zjištěny cíle, které se objevily na fixaci a zároveň úkol-irelevantní barva jednočetných těhotenství se blýskl v periferii. Do jaké míry úkol-irelevantní distraktorů v rozporu s centrální detekce úkol byl pak použit jako měřítko zdola nahoru pozornostní zachytit (Slanina a Egeth, 1994; Folk et al., 2002).
předpovědi byly následující: pacienti s lézí vynikající částí parietálního laloku (postihují SPL) by měl být narušena v top-down pozornost, orientace v úkolu (se zachovanou výkon na zachycení úkol), zatímco u pacientů s lézí v dolní části parietálního laloku (postihují TPJ), by měla být snížena na zachycení úkol (s ušetřen výkon na top-down úkol). Dvojí disociace této podobě nejen svědčí o nezávislé komponenty pozornost, ale také naznačuje, že tyto pozornostní komponenty jsou zprostředkovány nezávislé nervové mechanismy. Do studie bylo přijato osm pacientů s visuo-prostorovým zanedbáním a dokončili dva úkoly, klepnutím na stimulační nebo cílené zaměření pozornosti. Na základě jejich behaviorálního profilu byli pacienti rozděleni do skupin a jejich překrytí lézí bylo prozkoumáno (obrázek 2A). Pacienti, kteří vykazovali problémy s cílená pozornost, orientace, tak jak jsou kvantifikovány pomocí top-down pozornostní index (Obrázek 2B), prezentovány s lézí překrývají střed nad superior části parietální lalůček (subsuming SPL) s ušetřen nižší parietální lalůček (TPJ). Pacienti s lézí překrývají uprostřed dolní části parietální lalůček (subsuming TPJ), ale ušetřil SPL provádí obvykle na cílené orientace v úkolu, zatímco zbývající imunní vůči pozornostní zachytit (viz Obrázek 2C). Zjištění z této studie jasně naznačují, že SPL a TPJ jsou anatomické regiony, které jsou nutně přijati pro účely shora-dolů a zdola-nahoru orientace a že poškození SPL a TPJ vede k poruchám shora-dolů a zdola-nahoru orientace, resp.
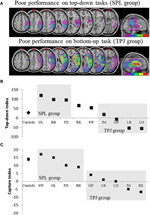
Obrázek 2. Výsledky neuropsychologické studie zaměřené na zkoumání relativního příspěvku SPL a TPJ k orientaci shora dolů a zdola nahoru. (A) Léze překrývá (fialová minimální přesah; červená maximální přesah) u pacientů, seskupených podle behaviorální deficity v top-down pozornost, orientace, označené SPL group (horní panel); a pacientů seskupeny podle behaviorální deficity v bottom-up orientace, označené TPJ skupiny (dolní panel). (B) behaviorální výkon na úkolu shora dolů shrnutý „indexem shora dolů“, který kvantifikuje rozdíly mezi prostorovými posuny shora dolů provedenými zleva doprava a naopak. Kontroly a skupina s lézemi TPJ vykazují podobnou účinnost při provádění prostorových posunů, zatímco pacienti s lézemi SPL vykazují sníženou účinnost. Skupinová kontrola a údaje o jednotlivých pacientech (označené iniciálami pacienta) jsou vyneseny na úsečku. (C) „Capture index“ je měřítkem bottom-up pozornost a kvantifikuje, do jaké míry úkol-irelevantní distraktorů zachytit pozornost od úkolu. Ovládací prvky a SPL léze skupina vykazují podobné hodnoty zachycení, tak, že obě skupiny jsou zachyceny úkol-irelevantní distraktory. TPJ léze skupina vykazují mnohem snížený index zachycení (selhání být zachycen). Vědomí, že pacienti byli umístěni v SPL nebo TPJ skupinu na základě chování, spíše než na základě léze, tedy vědomí, důslednost, s jakou pacienti končí v odpovídající skupině.
Interakce Mezi Top-Down a Bottom-Up Výběr
i když je zřejmě silná asociace mezi cílená orientace a SPL a stimul-řízený orientace a TPJ, data z Shomstein et al. (2010) studie pacientů naznačují, že tyto dva systémy nejsou zcela nezávislé. Tento závěr je podpořen zjištěním, že pacienti s poškozením SPL vykazovali vzorec výkonu označený jako “ hyper capture.“Na rozdíl od kontroly, pro koho cílit pouze barevné distraktoru zachytil pozornost (což vede k nižší cílové přesnost), irelevantní barevné distraktorů také ukázala být rušivé pro pacienty s SPL léze. Kromě toho, vzhledem k tomu, že pro ovládání pozornosti byl zajat distraktorů pouze tehdy, když jsou před nástupem cíl, pro pacienty s SPL léze pozornost, byl dokonce zajat distraktorů, předloží zároveň s cílem. Tento vzorec výkonu lze vysvětlit následujícím rámcem: SPL je zodpovědný za vedení pozornosti shora dolů, které zahrnuje určení aspektů podnětů, které jsou relevantní pro úkol (např. hledání červeného cíle; Corbetta a Shulman, 2002; Serences et al., 2005). Tento attentional set pak omezuje TPJ, takové, že zachycení pozornosti mechanismus, který je zprostředkován TPJ je pouze vyvolána úkol relevantní informace (např. červená distraktorů, že zachytí pozornost, a šedé distraktorů není zachytit pozornost při hledání červený terč). Absence SPL zabraňuje vytvoření úkolu relevantní attentional set, a tak nějaký podnět, úkol relevantní, nebo ne, je považováno za důležité proto, že zachytí pozornost (např. úkol-irelevantní distraktoru zachytit pozornost na SPL skupina) bez rozdílu.
bylo navrženo, že SPL a TPJ by mohly interagovat alespoň jedním ze dvou možných způsobů. První možností je, že TPJ slouží jako varovný systém, který detekuje behaviorálně relevantní podněty, ale postrádá vysoké prostorové rozlišení, tedy když behaviorálně relevantní stimul je zjištěna jeho přesné umístění, je dodávána v SPL, který ukládá prostorové mapy (Kastner et al., 1999; Wojciulik a Kanwisher, 1999; Bisley a Goldberg, 2003; Silver a kol., 2005). Související hypotetickou možností je, že zachycení mechanismu (to zahrnuje TPJ) působí jako jistič probíhající kognitivní aktivity při behaviorálně relevantní stimul je prezentována (Corbetta a Shulman, 2002, 2011). „Hyper-capture“ vzor aktivita pozorována u pacientů se zachovanou TPJ ale lesioned SPL poskytuje další důkazy pro hypotézu, že názory TPJ jako vydávání kontrolní signál, který ukončí úkol, na dosah ruky a tak slouží jako jistič (Corbetta a Shulman, 2002; Serences et al., 2005). Další nedávné neurozobrazování studie využívající různé vzory poskytly další důkazy pro interaktivní vztah mezi top-down a bottom-up, pozornost, orientace, a následně pro vztah mezi SPL a TPJ (Serences et al., 2005; Asplund et al., 2010; Diquattro a Geng, 2011).
Zatímco důkazy pro interakci mezi dva pozornostní systémy a dva pozornostní substráty (SPL a TPJ) je silný, to, co zůstává nejasné, je, zda tato interakce je přímý mezi SPL a TPJ, nebo zda to je provedeno prostřednictvím jiných přechodových regionů. Jak již bylo zmíněno, top-down a bottom-up, pozornost, orientace v sítích zapojit různé regiony v rámci frontální mozkové kůry, a proto je rozumné předpokládat, že konvergence mezi dvěma systémy může být provedeno pomocí čelního laloku. Dvě nedávné studie zkoumající interakce mezi top-down a bottom-up pozornostní výběr poskytla důkazy pro IFJ a IFG, jak je to možné stránek konvergence mezi stimul-řízené a cílené výběr (Asplund et al., 2010; Diquattro a Geng, 2011). V IFJ a IFG zdají být ideálními kandidáty pro takovou interakci vzhledem k jejich obecné zapojení v pozornosti a kognitivní kontrolou, stejně jako jeho účast v obou prostorových a non-prostorové výběr (Koechlin et al., 2003; Brass et al., 2005).
Role parietálního laloku při výběru shora dolů a zdola nahoru: Fyziologie
Zatímco důraz této recenze byly převážně umístěny na lidské studií, velké množství poznatků o zapojení parietální kůry v pozornostní orientace má být sbíráno z opice fyziologie šetření (viz nedávný přezkum Bisley a Goldberg, 2010). Nicméně, když přijde na zkoumání relativní příspěvky z různých oblastí do parietální kůry top-down a bottom-up, pozornost, orientace, opice, fyziologie literatura zaostává. Hlavním důvodem je to, že uvnitř opičí kůry se nezdá být důkaz pro stejnou segregaci kontroly shora dolů a zdola nahoru. Místo toho, boční intraparietal oblasti (RTU) původně myšlenka být zapojen v saccade plánování (Gnadt a Andersen, 1988) se podílí na vizuální pozornost a působí jako prioritní mapu, ve které vnější podněty jsou zastoupeny podle jejich chování prioritou odvozen buď shora dolů nebo zdola nahoru (Colby a Goldberg, 1999; Bisley a Goldberg, 2003, 2010; Balan a Gottlieb, 2006; Ipata et al., 2006; Buschman a Miller, 2007; Gottlieb a Balan, 2010).
Závěr
i když mnohem méně je známo o lidské temenní lalok než homologní opičí kůra, nedávné studie, využívající neuroimaging a neuropsychologické metody, začaly objasnění stále více jemnozrnné funkční a strukturální rozdíly. Tento přezkum se zaměřil na nedávné neuroimaging a neuropsychologické studie objasňují kognitivní role dorzální a ventrální oblasti parietální kůry v top-down a bottom-up, pozornost, orientace, a na interakce mezi dva pozornostní mechanismy přidělování.
Prohlášení o střetu zájmů
autor prohlašuje, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly být vykládány jako potenciální střet zájmů.
poděkování
tato práce byla podpořena grantem National Institutes of Health EY021644.
Asplund, C. L., Todd, J. J., Snyder, a. P., and Marois, R. (2010). Ústřední role laterální prefrontální kůry v cílené a stimulační pozornosti. Adresa. Neurovědci. 13, 507–512.
Pubmed Abstract | Pubmed Full Text / CrossRef Full Text
Bacon, W. F., and Egeth, h. E. (1994). Převažující zachycení pozornosti řízené stimulem. Vjem. Psychofyzik. 55, 485–496.
Pubmed Abstrakt / Pubmed Plný Text
Balan, P. F., a Gottlieb, J. (2006). Integrace exogenního vstupu do dynamické mapy salience odhalené rušivou pozorností. J. 26, 9239–9249.
Pubmed Abstraktní | Pubmed Plný Text | CrossRef Úplné Znění
Behrmann, M., Geng, j. J., a Shomstein, S. (2004). Parietální kůra a pozornost. Curre. Opine. Neurobiol. 14, 212–217.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Bisley, J.W., and Goldberg, M. E. (2003). Neuronální aktivita v laterální intraparietální oblasti a prostorová pozornost. Věda 299, 81-86.
Pubmed Abstraktní | Pubmed Plný Text | CrossRef Úplné Znění
Bisley, J. W., a Goldberg, M. E. (2010). Pozornost, záměr a priorita v parietálním laloku. Annu. Reverend Neurosci. 33, 1–21.
Pubmed Abstraktní | Pubmed Plný Text | CrossRef Úplné Znění
Brass, M., Derrfuss, J., Forstmann, B., a von Cramon, D. Y. (2005). Úloha dolní oblasti frontálního spojení v kognitivní kontrole. Trendy Cogn. Věda. 9, 314–316.
Pubmed Abstraktní | Pubmed Plný Text | CrossRef Úplné Znění
Buschman, T. J. Miller, E. K. (2007). Kontrola pozornosti shora dolů versus zdola nahoru v prefrontální a zadní parietální kortice. Věda 315, 1860-1862.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Chiu, Y. C., and Yantis, s. (2009). Doménově nezávislý zdroj kognitivní kontroly pro sady úloh: posun prostorové pozornosti a pravidla kategorizace přepínání. J. 29, 3930–3938.
Pubmed Abstraktní | Pubmed Plný Text | CrossRef Úplné Znění
Colby, C. L. a Goldberg, M. E. (1999). Prostor a pozornost v parietální kůře. Annu. Reverend Neurosci. 22, 319–349.
Pubmed Abstraktní | Pubmed Plný Text | CrossRef Úplné Znění
Corbetta, M., Kincade, J. M., Ollinger, J. M., McAvoy, M. P., a Shulman, G. L. (2000). Dobrovolná orientace je oddělena od detekce cíle v lidské zadní parietální kůře. Adresa. Neurovědci. 3, 292–297.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Corbetta, m., Kincade, M. J., Lewis, C., Snyder, A. Z., and Sapir, a. (2005). Neurální báze a obnova deficitů prostorové pozornosti při prostorovém zanedbávání. Adresa. Neurovědci. 8, 1603–1610.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Corbetta, m., Patel, G., and Shulman, g. l. (2008). Systém přeorientování lidského mozku: od prostředí k teorii mysli. Neuron 58, 306-324.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., and Shulman, G. L. (2002). Kontrola pozornosti zaměřené na cíl a podněty v mozku. Adresa. Reverend Neurosci. 3, 201–215.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Corbetta, M., and Shulman, g. l. (2011). Sítě prostorového zanedbávání a pozornosti. Annu. Reverend Neurosci. 34, 569–599.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Critchley, m. (1953). Parietální Laloky. Londýn, Velká Británie: Hafner Press.
Pubmed Abstraktní | Pubmed Úplné Znění
Diquattro, N. E., a Geng, j. J. (2011). Kontextové znalosti konfigurují sítě kontroly pozornosti. J. 31, 18026–18035.
Pubmed Abstraktní | Pubmed Plný Text | CrossRef Úplné Znění
Downar, J., Crawley, a. P., Mikulis, D. J., Davis, K. D. (2001). Vliv relevance úkolu na kortikální odpověď na změny vizuálních a sluchových podnětů: studie fMRI související s událostí. Neuroimage 14, 1256-1267.
Pubmed Abstraktní | Pubmed Plný Text | CrossRef Úplné Znění
Downar, J., Crawley, a. P., Mikulis, D. J., Davis, K. D. (2002). Kortikální síť citlivá na stimulační význam v neutrálním behaviorálním kontextu napříč různými smyslovými modalitami. J. Neurofyziol. 87, 615–620.
Pubmed Abstract / Pubmed Plný Text
Drummond, L., and Shomstein, s. (2010). Objektová pozornost: posun nebo nejistota? Attene. Vjem. Psychofyzik. 72, 1743–1755.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Egeth, h.E., and Yantis, s. (1997). Vizuální pozornost: kontrola, reprezentace a časový průběh. Annu. Reverend Psychol. 48, 269–297.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Eriksen, C.W., and Hoffman, je (1972). Časové a prostorové charakteristiky selektivního kódování z vizuálních displejů. Vjem. Psychofyzik. 12, 201–204.
Folk, C. L., Leber, a. B., and Egeth, h. E. (2002). Donutil tě mrknout! Kontingentní zachycení pozornosti vytváří prostorové blikání. Vjem. Psychofyzik. 64, 741–753.
Pubmed Abstract / Pubmed Plný Text
Folk, C. L., Remington, R. W., and Johnston, J. C. (1992). Nedobrovolné skryté orientace závisí na Nastavení kontroly pozornosti. J.Exp. Psychol. Hučení. Vjem. Provést. 18, 1030–1044.
Pubmed Abstract / Pubmed Plný Text
Friedrich, F. J., Egly, R., Rafal, R. D., and Beck, D. (1998). Deficity prostorové pozornosti u lidí: srovnání nadřazených parietálních a temporálně-parietálních lézí. Neuropsychologie 12, 193-207.
Pubmed Abstract | Pubmed Plný Text
Geng, J. J., and Behrmann, m. (2002). Pravděpodobnost určení cílové polohy usnadňuje vizuální vyhledávání implicitně u normálních účastníků a pacientů s hemispatiálním zanedbáním. Psychol. věda. 13, 520–525.
Pubmed Abstract | Pubmed Plný Text
Geng, J. J., and Behrmann, m. (2005). Prostorová Pravděpodobnost jako pozornost ve vizuálním vyhledávání. Vjem. Psychofyzik. 67, 1252–1268.
Pubmed Abstract / Pubmed Plný Text
Geng, J. J., and Mangun, G. R. (2011). Aktivace pravé temporoparietální křižovatky výraznou kontextovou narážkou usnadňuje diskriminaci cílů. Neuroimage 54, 594-601.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Gibson, B.S., and Kelsey, E. M. (1998). Zachycení pozornosti řízené stimulem závisí na sadě pozornosti pro vizuální prvky displeje. J.Exp. Psychol. Hučení. Vjem. Provést. 24, 699–706.
Pubmed Abstraktní | Pubmed Úplné Znění
Giesbrecht, B., Woldorff, M. G. Song, a. W., a Mangun, G. R. (2003). Nervové mechanismy řízení shora dolů během prostorové a rysové pozornosti. Neuroimage 19, 496-512.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Gnadt, J.W., and Andersen, R. a. (1988). Aktivita motorického plánování související s pamětí v zadní parietální kůře makaka. Expo. Mozek Res. 70, 216-220.
Pubmed Abstract / Pubmed Plný Text
Gottlieb, J., and Balan, P. (2010). Pozornost jako rozhodnutí v informačním prostoru. Trendy Cogn. Věda. 14, 240–248.
Pubmed Abstraktní | Pubmed Plný Text | CrossRef Úplné Znění
Greenberg, A. S., Esterman, M., Wilson, D., Serences, J. T., a Yantis, S. (2010). Řízení prostorové a rysové pozornosti v frontoparietální kůře. J. 30, 14330–14339.
Pubmed Abstraktní | Pubmed Plný Text | CrossRef Úplné Znění
Hopfinger, J. B., Buonocore, M. H., a Mangun, G. R. (2000). Nervové mechanismy kontroly pozornosti shora dolů. Adresa. Neurovědci. 3, 284–291.
Pubmed Abstrakt | Pubmed Plný Text / CrossRef Plný Text
Ipata, A. E., Gee, a. L., Gottlieb, J., Bisley, J. W., and Goldberg, M. E. (2006). Reakce rtů na popout stimul jsou sníženy, pokud jsou zjevně ignorovány. Adresa. Neurovědci. 9, 1071–1076.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Johnston, W.A., and Dark, v. J. (1986). Selektivní Pozornost. Annu. Reverend Psychol. 37, 43–75.
Juola, J. F., Koshino, H., and Warner, C. B. (1995). Kompromisy mezi pozornými účinky prostorových podnětů a náhlými nástupy. Vjem. Psychofyzik. 57, 333–342.
Pubmed Abstract / Pubmed Plný Text
Kastner, s., Pinsk, m. a., De Weerd, P., Desimone, R., and Ungerleider, L. G. (1999). Zvýšená aktivita v lidské zrakové kůře během zaměřené pozornosti při absenci vizuální stimulace. Neuron 22, 751-761.
Pubmed Abstract | Pubmed Full Text / CrossRef Full Text
Koechlin, e., Ody, C., and Kouneiher, F. (2003). Architektura kognitivní kontroly v lidské prefrontální kůře. Věda 302, 1181-1185.
Pubmed Abstract | Pubmed Full Text / CrossRef Full Text
Koshino, h., Warner, C. B., and Juola, J. F. (1992). Relativní účinnost centrálních, periferních a náhlých podnětů ve vizuální pozornosti. Q. J.Exp. Psychol. 45, 609-631.
Pubmed Abstraktní | Pubmed Úplné Znění
Liu, T., Slotnick, S. D., Serences, J. T., a Yantis, S. (2003). Kortikální mechanismy kontroly pozornosti založené na funkcích. Cereb. Cortex 13, 1334-1343.
Pubmed Abstract | Pubmed Full Text / CrossRef Full Text
Marois, R., Leung, HC, and Gore, JC (2000). Stimulační přístup k objektové identitě a zpracování polohy v lidském mozku. Neuron 25, 717-728.
Pubmed Abstraktní | Pubmed Plný Text | CrossRef Úplné Znění
McFire, J., a Zangwill, O. L. (1960). Visuo-konstruktivní postižení spojené s lézemi levé mozkové hemisféry. Brain 82, 243-260.
Moore, C.M., and Egeth, h. (1998). Jak pozornost založená na funkcích ovlivňuje vizuální zpracování? J.Exp. Psychol. Hučení. Vjem. Provést. 24, 1296–1310.
Pubmed Abstract / Pubmed Plný Text
Moran, J., a Desimone, R. (1985). Selektivní pozornost brány vizuální zpracování v extrastriate kůry. Věda 229, 782-784.
Pubmed Abstraktní | Pubmed Plný Text | CrossRef Úplné Znění
O’Craven, K., Rosen, B. R., Kwong, K. K., Treisman, A., a Savoy, R. L. (1997). Dobrovolná pozornost moduluje aktivitu fMRI v lidském MT-MST. Neuron 18, 591-598.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Piercy, m. (1964). Účinky mozkových lézí na intelektuální funkci: přehled současných výzkumných trendů. Br. J. Psychiatrie 110, 310-352.
Pubmed Abstract / Pubmed Plný Text
Posner, M. I. (1980). Orientace pozornosti. Q. J.Exp. Psychol. 32, 3–25.
Pubmed Abstraktní | Pubmed Úplné Znění
Posner, M. I., Snyder, C. R. a Davidson, B. J. (1980). Pozornost a detekce signálů. J.Exp. Psychol. 109, 160–174.
Pubmed Abstraktní | Pubmed Úplné Znění
Posner, M. I., Walker, J. a., Friedrich, F. J., a Rafal, R. D. (1984). Účinky parietálního poranění na skryté zaměření pozornosti. J. 4, 1863–1874.
Pubmed Abstraktní | Pubmed Úplné Znění
Serences, J. T., Shomstein, S., Leber, a. B., Golay, X, Egeth, H. E. a Yantis, S. (2005). Koordinace dobrovolné a stimulační kontroly pozornosti v lidské kůře. Psychol. Věda. 16, 114–122.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Shaw, M.L. (1978). Model přidělování kapacity pro reakční dobu. J.Exp. Psychol. Hučení. Vjem. Provést. 4, 586–598.
Shomstein, s., and Behrmann, m. (2006). Kortikální systémy zprostředkující vizuální pozornost jak objektům, tak prostorovým lokacím. Proc. Natle. Acad. Věda. USA 103, 11387-11392.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Shomstein, s., Lee, J., and Behrmann, m. (2010). Pokyny pro pozornost shora dolů a zdola nahoru: zkoumání role dorzálních a ventrálních parietálních Kortik. Expo. Brain Res.206, 197-208.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Shomstein, s., and Yantis, s. (2004a). Configurální a kontextová prioritizace v objektově založené pozornosti. Psychone. Býk. Rev. 11, 247-253.
Pubmed Abstract / Pubmed Plný Text
Shomstein, s., and Yantis, s. (2004b). Kontrola pozornosti se mění mezi zrakem a konkurzem v lidské kůře. J. 24, 10702–10706.
Pubmed Abstract | Pubmed Full Text / CrossRef Full Text
Shomstein, s., and Yantis, s. (2006). Parietální kůra zprostředkovává dobrovolnou kontrolu prostorové a neprostorové sluchové pozornosti. J. 26, 435–439.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Silver, M. a., Ress, D., a Heeger, D.J. (2005). Topografické mapy vizuální prostorové pozornosti v lidské parietální kůře. J. Neurofyziol. 94, 1358–1371.
Pubmed Abstraktní | Pubmed Plný Text | CrossRef Úplné Znění
Tamber-Rosenau, B. J., Esterman, M., Chiu, Y.-C., a Yantis, S. (2011). Kortikální mechanismy kognitivní kontroly pro posun pozornosti ve vidění a pracovní paměti. J. Cogn. Neurovědci. 23, 2905–2919.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Theeuwes, J. (1991). Exogenní a endogenní kontrola pozornosti: účinek vizuálních onsetů a offsetů. Vjem. Psychofyzik. 49, 83–90.
Pubmed Abstract / Pubmed Plný Text
Vallar, G., and Perani, d. (1986). Anatomie jednostranného zanedbávání po lézích mrtvice na pravé hemisféře. Klinická / CT-skenovací korelační studie u člověka. Neuropsychologie 24, 609-622.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Wojciulik, E., and Kanwisher, N. (1999). Obecnost parietálního zapojení do vizuální pozornosti. Neuron 23, 747-764.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Yantis, s., and Jonides, J. (1984). Náhlé vizuální náznaky a selektivní pozornost: důkazy z vizuálního vyhledávání. J.Exp. Psychol. Hučení. Vjem. Provést. 10, 601–621.
Pubmed Abstract / Pubmed Plný Text
Yantis, s., and Jonides, J. (1990). Náhlé vizuální onsety a selektivní pozornost: dobrovolná versus automatická alokace. J.Exp. Psychol. Hučení. Vjem. Provést. 16, 121–134.
Pubmed Abstrakt/Pubmed Plný Text
Yantis, s., Schwarzbach, J., Serences, J. T., Carlson, R. L., Steinmetz, m. a., Pekar, J. J., and Courtney, S. M. (2002). Přechodná nervová aktivita v lidské parietální kůře během posunů prostorové pozornosti. Adresa. Neurovědci. 5, 995–1002.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Yantis, s., and Serences, J. T. (2003). Kortikální mechanismy prostorové a objektové kontroly pozornosti. Curre. Opine. Neurobiol. 13, 187–193.
Pubmed Abstrakt | Pubmed Plný Text / CrossRef Plný Text