introduktion
vellykket interaktion med vores sensoriske miljø kræver en indviklet balance mellem to Opmærksomme selektionsmekanismer-den fra top til bund og bund. På vej over til produktionsgangen i dit lokale supermarked med det mål at samle få nødvendige ingredienser til mangosalaten, engagerer implementering af frivilligt målrettet, eller ovenfra og ned, opmærksomhedssystem, så du aktivt søger efter alle de krævede ingredienser blandt de mange valg af produkter. Imidlertid, skulle du høre en ringetone på en mobiltelefon, det vil sandsynligvis fange din opmærksomhed og afbryde din søgning. En sådan afbrydelse sker i en bottom-up, eller stimulus-drevet, mode, hvorved en simpel opmærksomhed af stimulus, det faktum, at ringen er forskellig fra andre lyde i dit miljø, anser det værdig udvælgelse. Det beskrevne scenario understreger vigtigheden af målrettet og stimulusdrevet udvælgelse for adfærd og peger på en fin balance, der skal eksistere mellem de to opmærksomhedssystemer for at forhindre “tunnelsyn” på den ene side og fuldstændig manglende evne til at fokusere på den anden.
Top-ned og Bottom-up valg: Adfærd
flere årtier med adfærdsmæssig forskning er blevet dedikeret til at demonstrere, at fordelingen af opmærksomhed kan styres af observatørens intentioner såvel som af den fysiske stimulans opmærksomhed. Meget af adfærdsmæssige beviser for top-ned og bottom-up opmærksomhedsallokering er blevet gennemgået grundigt andre steder (Johnston and Dark, 1986; Egeth og Yantis, 1997). For at opsummere viser undersøgelser, der viser effekter af opmærksomhedskontrol ovenfra og ned, at opmærksomheden med succes kan tildeles rumlige placeringer, funktioner, objekter osv., efter tilstedeværelse af eksogene eller endogene signaler (Eriksen og Hoffman, 1972; Posner, 1980; Posner et al., 1980), eller forventninger, der enten er fastsat ved forudgående viden eller ved hjælp af stimulus (Shav, 1978; Moore og Egeth, 1998; Geng og Behrmann, 2002, 2005; Shomstein og Yantis, 2004a; Drummond og Shomstein, 2010). Bevis, der understøtter bottom-up opmærksomhedsallokering, har påberåbt sig forskellige opmærksomhedsfangstparadigmer, hvor deltagerne er involveret i en søgning ovenfra og ned, og deres opmærksomhed omdirigeres til de opgave-irrelevante stimuli, hvilket viser, at opmærksomheden fanges af funktionssingletoner (unik vare; Yantis og Jonides, 1990; USA, 1991; Folk et al., 2002) og bratte onsets (Yantis og Jonides, 1984; USA, 1991; Koshino et al., 1992; Juola et al., 1995).
mens de fleste tidlige undersøgelser koncentrerede sig om at demonstrere bevis for opmærksomhedsvalg fra top til bund og bund, skiftede de seneste undersøgelser deres fokus til at undersøge, hvordan de to opmærksomhedsvalgssystemer interagerer. Denne undersøgelseslinje er drevet af observationer, at man for effektivt at vælge opgaverelevante oplysninger (f.eks. ingredienser til salaten) aktivt skal hæmme den opgaverelevante information, der ellers ville aflede opmærksomheden væk fra den aktuelle opgave. Bagsiden af denne logik er, at jo mindre man er fokuseret på opgaverelateret information, jo mere fangst vil der opstå. Det er blevet vist eksperimentelt, at observatørens Opmærksomme tilstand forudsiger, hvilken type information, og i hvilket omfang, i sidste ende vil fange opmærksomhed (Folk et al., 1992, 2002; Bacon og Egeth, 1994; Gibson og Kelsey, 1998). For eksempel Folk et al. (2002) viste, at når man søger efter et rødt bogstav, vil en observatør lettere blive fanget af en irrelevant stimulus i periferien, hvis denne stimulus er rød eller matcher målskabelonen på en eller anden måde. Da observatørens top-ned-kontrolindstillinger er indstillet til at søge efter en rød funktion, vil enhver stimulus, der er rød, sandsynligvis fange opmærksomhed og potentielt forstyrre top-ned-kontrol. Med en fangstopgave kan opmærksomhedssøgningsstrategier således skelnes fra hinanden ved at variere lighedsniveauerne mellem målets og distraktorernes stimulusegenskaber. Jo mere ens målet er for distraktoren, jo vanskeligere er det for observatøren at undgå fangst.
Parietallobens rolle i Top-ned og bottom-Up valg: Neuroimaging
forskellige neuroimaging-teknikker leverede stærke beviser for involvering af parietalbark i top-ned og bottom-up orientering, med beviserne gennemgået i vid udstrækning andetsteds (Corbetta og Shulman, 2002, 2011; Behrmann et al., 2004). Det er blevet påvist, at områder, der oftest aktiveres efter top-ned-signaler for at tage sig af bestemte placeringer, funktioner eller genstande, er placeret langs de dorsale dele af parietalbarken. Sådanne områder inkluderer ringere parietal lobule (IPL), dorsomediale regioner kaldet superior parietal lobule (SPL) samt flere mediale regioner langs precuneus gyrus (Yantis et al., 2002; Giesbrecht et al., 2003; Liu et al., 2003; Yantis og Serences, 2003; Figur 1). Flere top-ned-opgaver har vist sig med succes at engagere dorsale regioner i parietalbarken, nemlig dem, der involverer rumlig (Kastner et al., 1999; Corbetta et al., 2000; Hopfinger et al., 2000; Shomstein og Behrmann, 2006; Chiu og Yantis, 2009; Greenberg et al., 2010) samt ikke-rumlige skift af opmærksomhed (Giesbrecht et al., 2003; Yantis og Serences, 2003; Shomstein og Yantis, 2004b, 2006; Tamber-Rosenau et al., 2011).
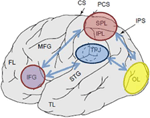
Figur 1. Skematisk skildring af relevante anatomiske landemærker projiceret på den laterale overflade af den menneskelige hjerne. Superior parietal lobule (SPL) og inferior parietal lobule (IPL) er regioner inden for den dorsale del af parietalbarken, der underordner opmærksomhedsorientering ovenfra og ned. Temporo-parietal junction (TPJ) er en region inden for den ventrale parietalbark, der underkaster sig opmærksomhed fra bunden opad. Begge, SPL og TPJ, menes at fremkalde styresignaler, der er ansvarlige for efterfølgende Opmærksomme moduleringer observeret over sensoriske regioner, i dette tilfælde modulerende (mærket med mørkeblå pile) visuelt fremkaldt aktivitet i occipital lap (OL). Derudover menes områder langs inferior frontal gyrus (IFG) og inferior frontal junction (IFJ) at tjene som konvergensområder for stimulusdrevet og oppefra og ned opmærksomhedskontrol (markeret med lyseblå tovejs pile).
i en typisk opgave, der sigter mod at engagere den opmærksomhedsallokering ovenfra og ned, vises enkeltpersoner to hurtige serielle visuelle præsentationsstrømme (RSVP) placeret perifert og instrueres oprindeligt om at overvåge en strøm til en kø (f.eks. et ciffer blandt strømmen af bogstaver). Cue ‘ s identitet angiver, om motivet skal bevare opmærksomheden på den aktuelle strøm eller skifte opmærksomhed til den anden strøm (Yantis et al., 2002; Yantis og Serences, 2003). To store fund observeres i sådanne paradigmer. Den første har at gøre med øget aktivering inden for de sensoriske regioner, der repræsenterer det aktuelle sted (f.eks. øget aktivitet inden for de venstre primære visuelle regioner, når den højre RSVP-strøm deltager). Dette fund giver fast bevis for, at deltagerne deltager i et bestemt sted, og at opmærksomheden modulerer styrken af det sensoriske respons (se figur 1; Moran og Desimone, 1985; O ‘ Craven et al., 1997). Det andet fund har at gøre med observationen om, at dorsale regioner i parietalloben selektivt aktiveres ved skift af top-ned-opmærksomhed. Det observeres, at SPL/IPL-aktivitetsforløbet er forbigående i naturen, hvilket antyder, at dette område af parietalbarken er kilden til et kort opmærksomt styresignal til at skifte Opmærksomme tilstande ovenfra og ned (Yantis et al., 2002).
flere fMRI-undersøgelser har dokumenteret, at bottom-up opmærksomhedsfangst, medieret af stimulus salience og/eller relevans, er underlagt temporo-parietal junction (TPJ; Figur 1). For eksempel, når emner deltager i og overvåger en ændring i enten en visuel eller auditiv stimulus, præsenteret samtidigt, forbedres aktivering af TPJ-regionerne i parietalloben. Ud over den tilsyneladende følsomhed over for relevante stimuli aktiveres TPJ også som reaktion på potentielt nye (uventede eller sjældne) begivenheder, når en organisme er involveret i en neutral adfærdsmæssig sammenhæng eller når den er involveret i en opgave (Marois et al., 2000; nedar et al., 2002; Serences et al., 2005; Corbetta et al., 2008; Asplund et al., 2010; Geng og Geng, 2011; Geng og Mangun, 2011). Denne aktivering sker uafhængigt af modaliteten (auditiv, taktil og visuel), hvor input leveres, hvilket afspejler TPJ ‘ s multisensoriske karakter (men se nedar et al., 2001).
i en typisk opgave, der undersøger den neurale mekanisme for bottom-up opmærksomhedsfangst, præsenteres deltagerne for en RSVP-strøm af genstande i midten af skærmen og bliver bedt om at identificere et foruddefineret mål (f.eks. identificere rødt bogstav præsenteret i en RSVP-strøm af hvide ikke-mål). En del af forsøgene indeholder en opgave-irrelevant fremtrædende distraktor præsenteret med forskellige tidsintervaller inden starten af målet, mens andre forsøg kun indeholder den fremtrædende distraktor (dvs.uden målet). “Target-distraktor” forsøg anvendes for at analysere omfanget af indfangning, hvilket viser, at den opgave-irrelevante distraktor faktisk er fremtrædende, hvilket giver et fald i målnøjagtighed. “Distraktor-in-isolation” – forsøgene bruges til yderligere analyser, da sådanne forsøg muliggør undersøgelse af aktivitet fremkaldt til den fremtrædende distraktor uden forurening fra de målrelaterede processer. Flere vigtige fund fremgår af sådanne paradigmer. For det første, når distraktorer er rumligt adskilt fra målplaceringen, ledsages fangstdistraktorer af øget kortikal aktivitet i tilsvarende regioner i sansebarken (f.eks. Sådanne resultater giver stærke beviser for, at rumlig opmærksomhed under indfangning faktisk fanges til den rumlige placering besat af distraktoren (Serences et al., 2005). For det andet aktiveres ventrale regioner i parietalbarken, hovedsageligt inden for TPJ, selektivt ved bottom-up, ufrivillig, skift af opmærksomhed. Ligesom aktivitet inden for SPL til orientering fra top til bund, aktivitetsforløbet observeret over TPJ er forbigående i naturen, hvilket antyder, at denne region er kilden til et kort opmærksomt styresignal for at skifte opmærksomhed på en bottom-up måde.
det skal bemærkes, at mens denne gennemgang er fokuseret på at adressere kognitive funktioner i den bageste parietalbark, rekrutteres andre regioner, især dem inden for frontalbarken, også til opmærksomhedsallokering fra top til bund og bund. Sådanne regioner inkluderer den ventrale frontale hjernebark (VFC), de frontale øjenfelter (FEF), inferior frontal junction (IFJ) og inferior frontal gyrus (IFG; Corbetta og Shulman, 2002, 2011; Serences et al., 2005; Asplund et al., 2010; Divattro og Geng, 2011).
Parietallobens rolle i Top-ned og bottom-Up valg: Neuropsykologi
Historisk stod forskere kritisk på neuropsykologiske undersøgelser af patienter med hemispatial forsømmelse (en forstyrrelse af rumlig tildeling af opmærksomhed til venstre hemi-rum) for at få indsigt i kognitive funktioner forbundet med parietallappen. I den klassiske neuropsykologiske litteratur blev parietalbarken som helhed generelt betragtet som det primære læsionssted for hemispatial forsømmelse. Denne opfattelse, uddybet i detaljer af tidlige forskere (Critchley, 1953; McFire og Sangvil, 1960; Piercy, 1964) anerkendte klart sammenhængen mellem parietal læsionen og den efterfølgende forsømmelse. Dette perspektiv blev stort set holdt gennem 1980 ‘ erne, da Posner og kolleger (1984) brugte det skjulte visuospatiale cueing-paradigme for at vise, at skader på parietallappen producerer et underskud i “frakobling” – operationen (tilbagetrækning af opmærksomhed fra et sted og skift til et andet), når målet er kontralateralt til læsionen. På trods af dette store fremskridt med at forstå det neurale grundlag for opmærksomhed og specifikt parietalbarkens “frakobling” – rolle antager deres fund imidlertid et enkelt kortikalt sted (parietalbark) og en enkelt funktionel kapacitet (“frakobling”). I modsætning til denne mere monolitiske tilgang til hjernen (parietalbark) og adfærd (opmærksom frigørelse) antyder det nylige adfærdsmæssige og neuroimaging-arbejde (gennemgået ovenfor og andre steder), at både den kortikale region og den tilhørende opmærksomhedsadfærd kan opdeles i kvalitativt forskellige profiler.
i betragtning af adskillelse af de kortikale netværk i ovenfra og ned og nedefra og op processer er en åbenbar forudsigelse, at skade på overlegne dele af parietal lobule (subsuming SPL) skulle give et underskud i målrettet opmærksomhedsorientering, mens skade på de underordnede dele af parietal lobule (subsuming TPJ) ville resultere i et underskud forbundet med stimulusdrevet opmærksomhedsfangst. I det omfang disse hjerneadfærdskorrespondancer er blevet undersøgt i den neuropsykologiske litteratur, opretholdes denne forudsigelse ikke åbenlyst. For eksempel er kliniske symptomer på hemispatial forsømmelse stærkt forbundet med skade på de underordnede dele af parietallappen, som inkluderer TPJ, snarere end til overlegne dele som SPL (Friedrich et al., 1998; Shomstein et al., 2010; Corbetta og Shulman, 2011). Dette er noget i strid med neuroimaging-litteraturen, hvilket antyder, at TPJ ‘ s rolle er i fangst af opmærksomhed snarere end i frivillig orientering af opmærksomhed, det domæne, hvor forsømmelsespatienter synes at have mest vanskeligheder. For at komplicere tingene yderligere er det blevet bemærket, at læsioner, der udelukkende involverer SPL, kun sjældent giver kliniske tegn på forsømmelse (Vallar og Perani, 1986). En anden nylig undersøgelse med patienter med læsioner centreret primært over TPJ og STG, men bevaret SPL, Corbetta et al. (2005) viste, at rumlig forsømmelse såvel som dens bedring var forbundet med gendannelse af aktivitet i både de ventrale temporo-parietale og dorsale parietale regioner (se Corbetta and Shulman, 2011 til en gennemgang). Mens det er interessant og spændende i sine konklusioner, adskiller denne sidste undersøgelse ikke det relative bidrag fra dorsale og ventrale veje til forskellige typer opmærksomhed, da patienter kun blev testet på en variant af Posner skjult rumlig opmærksomhedsopgave, opgave, der menes at engagere både oppefra og nedefter opmærksomhedsorientering.
for at skelne mellem måldrevet opmærksomhedskontrol og fremtrædende opmærksomhedsfangst og for at undersøge deres kortlægning på henholdsvis SPL og TPJ vedtog den nylige undersøgelse to adfærdsmæssige paradigmer, der hver var rettet mod en af disse former for opmærksomhed (Shomstein et al., 2010). For at undersøge integriteten af top-ned opmærksomhedsorientering hos patienterne blev der anvendt en top-ned-opgave, der krævede, at deltagerne skiftede rumlig opmærksomhed mellem de rumligt adskilte RSVP-strømme (en opgave, der med succes er blevet brugt til at demonstrere SPL-aktivering i fMRI-studier (Yantis et al., 2002)). Tilsvarende, for at undersøge patienternes bottom-up Opmærksomme orienterende evner, en variant af Folk et al. (2002) kontingent capture paradigme blev anvendt, hvor deltagerne opdagede mål, der dukkede op ved fiksering, mens opgave-irrelevante farvesingletoner blev blinket i periferien. I hvilket omfang opgave-irrelevante distraktorer forstyrrer den centrale detektionsopgave blev derefter brugt som et mål for bottom-up attentional capture (Bacon og Egeth, 1994; Folk et al., 2002).
forudsigelserne var som følger: patienter med læsioner til overlegne dele af parietallappen (påvirker SPL) bør være nedsat i den opmærksomhedsorienterende opgave fra top til bund (med bevaret ydeevne på fangstopgaven), mens patienter med læsioner til de ringere dele af parietallappen (påvirker TPJ) bør være nedsat på fangstopgaven (med sparet ydeevne på opgaven fra top til bund). En dobbelt dissociation af denne form vidner ikke kun om de uafhængige opmærksomhedskomponenter, men antyder også, at sådanne Opmærksomme komponenter formidles af uafhængige neurale mekanismer. Otte patienter med visuo-rumlig forsømmelse blev rekrutteret til undersøgelsen og afsluttede to opgaver og tappede enten stimulusdrevet eller målrettet opmærksomhedsorientering. Baseret på deres adfærdsprofil blev patienterne sorteret i grupper, og deres læsionsoverlapning blev undersøgt (figur 2a). Patienter, der udviste vanskeligheder med målrettet opmærksomhedsorientering, som kvantificeret af opmærksomhedsindekset ovenfra og ned (figur 2b), præsenteret med læsionsoverlapning centreret over overlegne dele af parietal lobule (subsuming SPL) med skånet ringere parietal lobule (TPJ). Patienter med læsionsoverlapning centreret over de ringere dele af parietal lobule (subsuming TPJ), men skånet SPL udført normalt på den målstyrede orienteringsopgave, mens de forbliver immun mod opmærksom fangst (figur 2C). Resultaterne fra denne undersøgelse tyder tydeligt på, at SPL og TPJ er anatomiske regioner, der nødvendigvis rekrutteres med henblik på top-ned og bottom-up orientering, og at skade på SPL og TPJ fører til forstyrrelser i henholdsvis top-ned og bottom-up orientering.
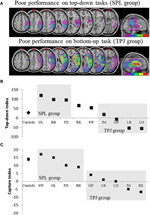
figur 2. Resultater af den neuropsykologiske undersøgelse med det formål at undersøge det relative bidrag fra SPL og TPJ til top-ned og bottom-up orientering. (A) Læsionsoverlapninger (lilla minimal overlapning; rød maksimal overlapning) for patienter grupperet efter adfærdsmæssige underskud i oppefra og ned opmærksomhedsorientering, mærket SPL-gruppen (øverste panel); og patienter grupperet efter adfærdsmæssige underskud i bottom-up orientering, mærket TPJ-gruppen (nederste panel). (B) adfærdsmæssig præstation på top-ned-opgaven opsummeret med et “top-ned-indeks”, der kvantificerer forskelle mellem rumlige top-ned-skift foretaget fra venstre mod højre og omvendt. Kontroller og den TPJ-læsionerede gruppe viser lignende effektivitet ved udførelse af rumlige skift, mens patienter med SPL-læsioner viser nedsat effektivitet. Gruppekontrol og individuelle patientdata (mærket med patientens initialer) er afbildet på abscissen. (C) “Capture indeks” er et mål for bottom-up opmærksomhed og kvantificerer i hvilket omfang opgave-irrelevante distraktorer fange opmærksomhed væk fra opgaven. Kontrolelementer og den SPL-læsionerede gruppe viser lignende fangstværdier, således at begge grupper fanges af de opgave-irrelevante distraktorer. TPJ lesioned group viser meget reduceret fangstindeks (manglende optagelse). Bemærk, at patienter blev placeret i SPL-eller TPJ-gruppen baseret på adfærd snarere end baseret på læsionen, og bemærk således konsistensen, som patienter ender med i den tilsvarende gruppe.
interaktion mellem Top-ned og bottom-up valg
selvom der tilsyneladende er en stærk sammenhæng mellem målrettet orientering og SPL og stimulusdrevet orientering og TPJ, data fra Shomstein et al. (2010) patientundersøgelse antyder, at disse to systemer ikke er helt uafhængige. Denne konklusion understøttes af konstateringen af, at patienter med SPL-skade udviste et præstationsmønster mærket som “hyper capture.”I modsætning til kontroller, for hvem kun målfarvet distraktor fangede opmærksomhed (hvilket førte til lavere målnøjagtighed), viste irrelevante farvede distraktorer sig også at være distraherende for patienter med SPL-læsion. Derudover, mens opmærksomheden for kontroller kun blev fanget af distraktorer, når de gik forud for starten af målet, for patienter med SPL-læsioner blev opmærksomheden endda fanget af distraktorer præsenteret samtidig med målet. Dette præstationsmønster kan forklares ved følgende rammer: SPL er ansvarlig for vejledning af opmærksomhed ovenfra og ned, der inkluderer bestemmelse af aspekterne af de stimuli, der er relevante for opgaven (f.eks. Søg efter rødt mål; Corbetta og Shulman, 2002; Serences et al., 2005). Dette opmærksomhedssæt begrænser derefter TPJ, således at indfangning af opmærksomhedsmekanisme, der formidles af TPJ, kun udløses af opgaven relevant information (f.eks. røde distraktorer fanger opmærksomhed, og grå distraktorer fanger ikke opmærksomhed, når de søger efter et rødt mål). Fraværet af SPL forhindrer etablering af en opgave relevant opmærksomhed sæt og dermed enhver stimulus, opgave relevant eller ej, anses for vigtigt derfor fange opmærksomhed (f.eks opgave-irrelevant distraktor fange opmærksomhed for SPL-gruppen) flæng.
det er blevet foreslået, at SPL og TPJ kunne interagere på mindst en af to mulige måder. Den første mulighed er, at TPJ fungerer som et alarmeringssystem, der registrerer adfærdsmæssigt relevante stimuli, men mangler den høje rumlige opløsning, således når en adfærdsmæssigt relevant stimulus opdages, leveres dens nøjagtige placering af SPL, der gemmer rumlige kort (Kastner et al., 1999, 1999, Bisley og Goldberg, 2003, Silver et al., 2005). En relateret hypotetisk mulighed er, at fangstmekanismen (der inkluderer TPJ) fungerer som en afbryder for løbende kognitiv aktivitet, når en adfærdsmæssigt relevant stimulus præsenteres (Corbetta og Shulman, 2002, 2011). Det “hyper-capture” aktivitetsmønster observeret hos patienter med konserveret TPJ, men læsioneret SPL, giver yderligere bevis for hypotesen, der betragter TPJ som et styresignal, der afslutter den aktuelle opgave og dermed tjener som en afbryder (Corbetta og Shulman, 2002; Serences et al., 2005). Andre nylige neuroimaging-undersøgelser, der anvender forskellige paradigmer, har givet yderligere bevis for et interaktivt forhold mellem top-ned og bottom-up opmærksomhedsorientering og derefter for forholdet mellem SPL og TPJ (Serences et al., 2005; Asplund et al., 2010; Geng, 2011).
mens beviset for en interaktion mellem de to opmærksomhedssystemer og de to opmærksomhedsunderlag (SPL og TPJ) er stærkt, er det uklart, om denne interaktion er direkte mellem SPL og TPJ, eller om den opnås gennem andre mellemliggende regioner. Som tidligere nævnt engagerer opmærksomhedsorienterende netværk fra top til bund forskellige regioner inden for frontalbarken, og det er derfor rimeligt at antage, at konvergensen mellem de to systemer kan opnås via frontallappen. To nylige undersøgelser, der undersøgte interaktionen mellem opmærksomhedsvalg fra top til bund og bund, fremlagde bevis for IFJ og IFG som mulige steder for konvergens mellem stimulusdrevet og målrettet udvælgelse (Asplund et al., 2010; Divattro og Geng, 2011). IFJ og IFG ser ud til at være ideelle kandidater til sådan interaktion i betragtning af deres generelle engagement i opmærksomhed og kognitiv kontrol såvel som dets engagement i både rumlig og ikke-rumlig udvælgelse (Koechlin et al., 2003; Brass et al., 2005).
Parietallobens rolle i Top-ned og bottom-Up valg: Fysiologi
mens vægten af denne gennemgang overvejende er lagt på menneskelige studier, er et stort væld af viden om involvering af parietalbark I opmærksom orientering hentet fra abefysiologiundersøgelser (Se nylig gennemgang af Bisley and Goldberg, 2010). Imidlertid, når det kommer til at undersøge de relative bidrag fra forskellige regioner inden for parietalbarken til opmærksomhedsorientering fra top til bund og bund, abe fysiologi litteratur kommer til kort. Den primære årsag til dette er, at der inden for abebarken ikke synes at være tegn på den samme adskillelse af top-ned og bottom-up kontrol. I stedet menes lateral intraparietal område (læbe) oprindeligt at være involveret i saccadeplanlægning (Gnadt og Andersen, 1988) er involveret i visuel opmærksomhed og fungerer som et prioritetskort, hvor eksterne stimuli er repræsenteret i henhold til deres adfærdsmæssige prioritet afledt enten ovenfra og ned eller nedefra og op (Colby og Goldberg, 1999; Bisley og Goldberg, 2003, 2010; Balan og Gottlieb, 2006; Ipata et al., 2006; Buschman og Miller, 2007; Gottlieb og Balan, 2010).
konklusion
selvom der er meget mindre kendt om human parietalbark end for homolog abebark, er nylige undersøgelser, der anvender neuroimaging og neuropsykologiske metoder, begyndt at belyse stadig finkornede funktionelle og strukturelle forskelle. Denne gennemgang fokuserede på nylige neuroimaging og neuropsykologiske undersøgelser, der belyser de kognitive roller i dorsale og ventrale regioner i parietalbarken i opmærksomhedsorientering fra top til bund og bund og på interaktionen mellem de to opmærksomhedsallokeringsmekanismer.
interessekonflikt Erklæring
forfatteren erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
anerkendelser
dette arbejde blev støttet af National Institutes of Health grant EY021644.
Asplund, C. L., Todd, J. J., Snyder, A. P. og Marois, R. (2010). En central rolle for den laterale præfrontale hjernebark i målrettet og stimulusdrevet opmærksomhed. Nat. Neurosci. 13, 507–512.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Bacon, F. F. og Egeth, H. E. (1994). Altoverskyggende stimulus-drevet opmærksomhedsfangst. Percept. Psykofys. 55, 485–496.
Pubmed abstrakt/Pubmed Fuld tekst
Balan, P. F., og Gottlieb, J. (2006). Integration af eksogent input i et dynamisk salience-kort afsløret af forstyrrende opmærksomhed. J. Neurosci. 26, 9239–9249.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Behrmann, M., Geng, J. J. og Shomstein, S. (2004). Parietal bark og opmærksomhed. Curr. Opin. Neurobiol. 14, 212–217.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Bisley, J. V. og Goldberg, M. E. (2003). Neuronal aktivitet i det laterale intraparietale område og rumlig opmærksomhed. Videnskab 299, 81-86.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Bisley, J. V. og Goldberg, M. E. (2010). Opmærksomhed, intention og prioritet i parietallappen. Annu. Pastor Neurosci. 33, 1–21.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
messing, M., Derrfuss, J., Forstmann, B. Og von Cramon, D. Y. (2005). Rollen af det underordnede frontale knudeområde i kognitiv kontrol. Tendenser Cogn. Sci. 9, 314–316.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Buschman, T. J. og Miller, E. K. (2007). Top-ned versus bottom-up kontrol af opmærksomhed i præfrontale og posterior parietale cortices. Videnskab 315, 1860-1862.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Chiu, Y. C. og Yantis, S. (2009). En domæne-uafhængig kilde til kognitiv kontrol for opgavesæt: skiftende rumlig opmærksomhed og skifte kategoriseringsregler. J. Neurosci. 29, 3930–3938.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Colby, C. L. og Goldberg, M. E. (1999). Plads og opmærksomhed i parietalbarken. Annu. Pastor Neurosci. 22, 319–349.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Corbetta, M., Kincade, J. M., Ollinger, J. M., McAvoy, M. P. og Shulman, G. L. (2000). Frivillig orientering adskilles fra måldetektion i human posterior parietalbark. Nat. Neurosci. 3, 292–297.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Corbetta, M., Kincade, M. J., C., Snyder, A. Å, og Sapir, A. (2005). Neuralt grundlag og genopretning af rumlige opmærksomhedsunderskud i rumlig forsømmelse. Nat. Neurosci. 8, 1603–1610.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Corbetta, M., Patel, G. og Shulman, G. L. (2008). Det omorienterende system i den menneskelige hjerne: fra miljø til sindsteori. Neuron 58, 306-324.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Corbetta, M., Og Shulman, G. L. (2002). Kontrol af målrettet og stimulusdrevet opmærksomhed i hjernen. Nat. Pastor Neurosci. 3, 201–215.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Corbetta, M., and Shulman, G. L. (2011). Spatial forsømmelse og opmærksomhed netværk. Annu. Pastor Neurosci. 34, 569–599.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Critchley, M. (1953). Parietalloberne. London, Storbritannien: Hafner Press.
Pubmed abstrakt/Pubmed Fuld tekst
Divatro, N. E. og Geng, J. J. (2011). Kontekstuel viden konfigurerer opmærksomhedsstyringsnetværk. J. Neurosci. 31, 18026–18035.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
nedar, J., A. P., Mikulis, D. J. og Davis, K. D. (2001). Effekten af opgaverelevans på det kortikale respons på ændringer i visuel og auditiv stimuli: en hændelsesrelateret fMRI-undersøgelse. Neurobillede 14, 1256-1267.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
nedar, J., A. P., Mikulis, D. J. og Davis, K. D. (2002). Et kortikalt netværk, der er følsomt over for stimulus salience i en neutral adfærdsmæssig sammenhæng på tværs af flere sensoriske modaliteter. J. Neurofysiol. 87, 615–620.
Pubmed abstrakt/Pubmed Fuld tekst
Drummond, L. og Shomstein, S. (2010). Objektbaseret opmærksomhed: skift eller usikkerhed? Atten. Percept. Psykofys. 72, 1743–1755.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Egeth, H. E. og Yantis, S. (1997). Visuel opmærksomhed: kontrol, repræsentation og tidsforløb. Annu. Pastor Psychol. 48, 269–297.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Eriksen, C. V. og Hoffman, J. E. (1972). Tidsmæssige og rumlige egenskaber ved selektiv kodning fra visuelle skærme. Percept. Psykofys. 12, 201–204.
Folk, C. L., Leber, A. B. Og Egeth, H. E. (2002). Fik dig til at blinke! Betinget opmærksomhedsfangst producerer et rumligt blink. Percept. Psykofys. 64, 741–753.
Pubmed abstrakt/Pubmed Fuld tekst
Folk, C. L., Remington, R. V. og Johnston, J. C. (1992). Ufrivillig skjult orientering er betinget af indstillinger for opmærksomhedskontrol. J. Eksp. Psychol. Hum. Percept. Udføre. 18, 1030–1044.
Pubmed abstrakt/Pubmed Fuld tekst
Friedrich, F. J., Egly, R., Rafal, R. D. og Beck, D. (1998). Rumlige opmærksomhedsunderskud hos mennesker: en sammenligning af overlegne parietale og tidsmæssige parietale knudelæsioner. Neuropsykologi 12, 193-207.
Pubmed abstrakt/Pubmed Fuld tekst
Geng, J. J. og Behrmann, M. (2002). Sandsynlighed cuing af målplacering Letter visuel søgning implicit hos normale deltagere og patienter med hemispatial forsømmelse. Psychol. sci. 13, 520–525.
Pubmed abstrakt/Pubmed Fuld tekst
Geng, J. J. og Behrmann, M. (2005). Rumlig sandsynlighed som en opmærksom cue i visuel søgning. Percept. Psykofys. 67, 1252–1268.
Pubmed abstrakt/Pubmed Fuld tekst
Geng, J. J. og Mangun, G. R. (2011). Højre temporoparietal junction aktivering af en fremtrædende kontekstuel cue Letter måldiskrimination. Neuroimage 54, 594-601.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Gibson, B. S. og Kelsey, E. M. (1998). Stimulusdrevet opmærksomhedsfangst er betinget af opmærksomhedssæt til displaybrede visuelle funktioner. J. Eksp. Psychol. Hum. Percept. Udføre. 24, 699–706.
Pubmed abstrakt/Pubmed Fuld tekst
Giesbrecht, B., Uldorff, M. G., sang, A. V. og Mangun, G. R. (2003). Neurale mekanismer for top-ned kontrol under rumlig og funktion opmærksomhed. Neuroimage 19, 496-512.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Gnadt, J. V. og Andersen, R. A. (1988). Hukommelsesrelateret motorplanlægningsaktivitet i posterior parietalbark af makak. Eksp. Brain Res. 70, 216-220.
Pubmed abstrakt/Pubmed Fuld tekst
Gottlieb, J. og Balan, P. (2010). Opmærksomhed som en beslutning i informationsrummet. Tendenser Cogn. Sci. 14, 240–248.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Greenberg, A. S., Esterman, M., D., Serences, J. T. og Yantis, S. (2010). Kontrol af rumlig og funktionsbaseret opmærksomhed i frontoparietalbarken. J. Neurosci. 30, 14330–14339.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Hopfinger, J. B., Buonocore, M. H. og Mangun, G. R. (2000). De neurale mekanismer til top-ned opmærksomhedskontrol. Nat. Neurosci. 3, 284–291.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Ipata, A. E., Gee, A. L., Gottlieb, J., Bisley, J. V. og Goldberg, M. E. (2006). LIP reaktioner på en popout stimulus reduceres, hvis det er åbenlyst ignoreret. Nat. Neurosci. 9, 1071–1076.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Johnston, V. J. (1986). Selektiv Opmærksomhed. Annu. Pastor Psychol. 37, 43–75.
Juola, J. F., Koshino, H. og Varner, C. B. (1995). Afvejninger mellem Opmærksomme effekter af rumlige signaler og pludselige ændringer. Percept. Psykofys. 57, 333–342.
Pubmed abstrakt/Pubmed Fuld tekst
Kastner, S., Pinsk, M. A., De Vierd, P., Desimone, R. og Ungerleider, L. G. (1999). Øget aktivitet i menneskelig visuel bark under rettet opmærksomhed i fravær af visuel stimulering. Neuron 22, 751-761.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Koechlin, E., Ody, C. og Kouneiher, F. (2003). Arkitekturen for kognitiv kontrol i den menneskelige præfrontale hjernebark. Videnskab 302, 1181-1185.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Koshino, H. C. B. Og Juola, J. F. (1992). Relativ effektivitet af centrale, perifere og pludselige tegn i visuel opmærksomhed. K. J. Eksp. Psychol. En 45, 609-631.
Pubmed abstrakt/Pubmed Fuld tekst
Liu, T., Slotnick, S. D., Serences, J. T. og Yantis, S. (2003). Kortikale mekanismer til funktionsbaseret opmærksomhedskontrol. Cereb. Bark 13, 1334-1343.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Marois, R., Leung, H. C. og Gore, J. C. (2000). En stimulusdrevet tilgang til objektidentitet og lokalitetsbehandling i den menneskelige hjerne. Neuron 25, 717-728.
Pubmed Abstract / Pubmed Fuld tekst / CrossRef Fuld tekst
McFire, J., og Sangvil, O. L. (1960). Visuo-konstruktive handicap forbundet med læsioner i venstre hjernehalvdel. Hjerne 82, 243-260.
Moore, C. M. Og Egeth, H. (1998). Hvordan påvirker funktionsbaseret opmærksomhed visuel behandling? J. Eksp. Psychol. Hum. Percept. Udføre. 24, 1296–1310.
Pubmed abstrakt / Pubmed Fuld tekst
Moran, J. Desimone, R. (1985). Selektiv opmærksomhed gates visuel behandling i ekstrastriatbarken. Videnskab 229, 782-784.
Pubmed abstrakt | Pubmed Fuld tekst/CrossRef Fuld tekst
O ‘ Craven, K., Rosen, B. R., K. K., Treisman, A. og Savoy, R. L. (1997). Frivillig opmærksomhed modulerer fMRI-aktivitet i menneskelig MT-MST. Neuron 18, 591-598.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Piercy, M. (1964). Virkningerne af cerebrale læsioner på intellektuel funktion: en gennemgang af aktuelle forskningstendenser. Br. J. Psykiatri 110, 310-352.
Pubmed abstrakt/Pubmed Fuld tekst
Posner, M. I. (1980). Orientering af opmærksomhed. K. J. Eksp. Psychol. 32, 3–25.
Pubmed abstrakt/Pubmed Fuld tekst
Posner, M. I., Snyder, C. R. og Davidson, B. J. (1980). Opmærksomhed og påvisning af signaler. J. Eksp. Psychol. 109, 160–174.
Pubmed abstrakt/Pubmed Fuld tekst
Posner, M. I., rollator, J. A., Friedrich, F. J. og Rafal, R. D. (1984). Virkninger af parietal skade på skjult orientering af opmærksomhed. J. Neurosci. 4, 1863–1874.
Pubmed abstrakt/Pubmed Fuld tekst
Serences, J. T., Shomstein, S., Leber, A. B., Golay, Egeth, H. E. og Yantis, S. (2005). Koordinering af frivillig og stimulusdrevet opmærksomhedskontrol i menneskelig hjernebark. Psychol. Sci. 16, 114–122.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Shav, M. L. (1978). En kapacitetstildelingsmodel for reaktionstid. J. Eksp. Psychol. Hum. Percept. Udføre. 4, 586–598.
Shomstein, S., og Behrmann, M. (2006). Kortikale systemer, der formidler visuel opmærksomhed på både objekter og rumlige placeringer. Proc. Natl. Acad. Sci. U. S. A. 103, 11387-11392.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Shomstein, S., Lee, J. og Behrmann, M. (2010). Top-ned og bottom-up opmærksomhedsvejledning: undersøgelse af rollen af dorsale og ventrale parietale cortices. Eksp. Brain Res. 206, 197-208.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Shomstein, S. og Yantis, S. (2004a). Konfiguration og kontekstuel prioritering i objektbaseret opmærksomhed. Psychon. Bull. Åb 11, 247-253.
Pubmed abstrakt/Pubmed Fuld tekst
Shomstein, S. og Yantis, S. (2004b). Kontrol af opmærksomhed skifter mellem vision og audition i menneskelig hjernebark. J. Neurosci. 24, 10702–10706.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Shomstein, S., og Yantis, S. (2006). Parietalbarken formidler frivillig kontrol af rumlig og ikke-rumlig auditiv opmærksomhed. J. Neurosci. 26, 435–439.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Sølv, M. A., Ress, D., og Heeger, D. J. (2005). Topografiske kort over visuel rumlig opmærksomhed i menneskelig parietalbark. J. Neurofysiol. 94, 1358–1371.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Tamber-Rosenau, B. J., Esterman, M., Chiu, Y.-C. og Yantis, S. (2011). Kortikale mekanismer for kognitiv kontrol til at skifte opmærksomhed i syn og arbejdshukommelse. J. Cogn. Neurosci. 23, 2905–2919.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Theeuves, J. (1991). Eksogen og endogen kontrol af opmærksomhed: effekten af visuelle onsets og offsets. Percept. Psykofys. 49, 83–90.
Pubmed abstrakt/Pubmed Fuld tekst
Vallar, G. og Perani, D. (1986). Anatomien af ensidig forsømmelse efter højre hjernehalvdel slagtilfælde læsioner. En klinisk / CT-scan korrelationsundersøgelse hos mennesker. Neuropsykologi 24, 609-622.
Pubmed abstrakt | Pubmed Fuld tekst/CrossRef Fuld tekst
PubMed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
PubMed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Generaliteten af parietal involvering i visuel opmærksomhed. Neuron 23, 747-764.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Yantis, S., og Jonides, J. (1984). Pludselige visuelle påvirkninger og selektiv opmærksomhed: bevis fra visuel søgning. J. Eksp. Psychol. Hum. Percept. Udføre. 10, 601–621.
Pubmed abstrakt/Pubmed Fuld tekst
Yantis, S. og Jonides, J. (1990). Pludselige visuelle påvirkninger og selektiv opmærksomhed: frivillig versus automatisk tildeling. J. Eksp. Psychol. Hum. Percept. Udføre. 16, 121–134.
PubMed Abstract/Pubmed Fuld tekst
Yantis, S. Carlson, R. L., Steinmetts, M. A., Pekar, J. J. og Courtney, S. M. (2002). Transient neural aktivitet i human parietalbark under rumlige opmærksomhedsskift. Nat. Neurosci. 5, 995–1002.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst
Yantis, S., and Serences, J. T. (2003). Kortikale mekanismer for rumbaseret og objektbaseret opmærksomhedskontrol. Curr. Opin. Neurobiol. 13, 187–193.
Pubmed abstrakt / Pubmed Fuld tekst / CrossRef Fuld tekst