introduktion
fossile ressourcer er det vigtigste råmateriale til energi og organiske forbindelser, og deres anvendelse resulterer i emission af drivhusgasser forbundet med klimaændringer. Det kommende klimaulykke kræver en presserende overgang fra en fossil-baseret til en biobaseret økonomi, hvor lignocellulosisk biomasse snarere end olie bruges til produktion af brændstoffer, kemikalier og materialer. Træ er en vigtig kilde til lignocellulosisk biomasse; det består hovedsageligt af sekundære fortykkede cellevægge rig på cellulose, hemicelluloser og lignin. Alle tre polymerer kan valoriseres i den biobaserede Økonomi. Cellulose er en kilde til papirmasse-og papirindustrien, og både cellulose og hemicelluloser kan depolymeriseres til deres monosaccharider til gæring til f.eks. bioethanol, mælkesyre og vaskemidler (Vanholme et al., 2013b). Da lignin negativt påvirker effektiviteten af træforarbejdning mod disse applikationer, kan træer konstrueres til at akkumulere mindre lignin for at blive mere modtagelige for produktion af papir og gærbare sukkerarter. På den anden side betragtes lignin i stigende grad som en værdifuld komponent i den biobaserede Økonomi. I betragtning af at lignin er den største vedvarende aromatiske kilde på jorden, kan et bioraffinaderis økonomiske levedygtighed øges markant, hvis lignin også værdiseres og bruges som en ressource til produktion af kemikalier (Holladay et al., 2007; Tuck et al., 2012; Davis et al., 2013; Ragauskas et al., 2014; Li C. Et Al., 2015; Van Den Bosch et al., 2015; Rinaldi et al., 2016; Upton og Kasko, 2016; Schutyser et al., 2018).ligninpolymeren er sammensat af monolignoler, der produceres af de biosyntetiske phenylpropanoid-og monolignolveje ved en række reaktioner, der begynder med deaminering af phenylalanin (Figur 1). Det er en af de mest almindelige årsager til denne sygdom, som er forårsaget af en række sygdomme, der kan føre til en alvorlig sygdom., 2011; Jens et al., 2013). Disse monolignolradikaler parrer sig derefter med hinanden på en kombinatorisk måde og genererer en række kemiske bindinger, såsom aryl-etherbinding (kurr-o-4), resinolbinding (kurr-Karr) og phenylcoumaranbinding (kurr-5) (Boerjan et al., 2003; Ralph et al., 2004; Vanholme et al., 2010). De mest almindelige monolignoler er p-coumaryl -, coniferyl-og sinapylalkoholer, som genererer H -, G-og S-enhederne ved deres inkorporering i ligninpolymeren (2010; Ralph et al., 2019; Vanholme et al., 2019). Lignin fra nåletræ (gymnospermer) består næsten udelukkende af G-enheder med en mindre brøkdel af H-enheder, mens lignin fra hårdttræ (angiospermer) har s-enheder ud over G-enheder og spor af H-enheder (Boerjan et al., 2003; Vanholme et al., 2010, 2019). Udover disse traditionelle monolignoler kan en række andre p-hydroksylerede aromatiske molekyler inkorporeres i ligninpolymeren til forskellige niveauer (Vanholme et al., 2019).
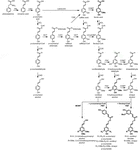
Figure 1. Lignin biosynthetic pathway. Alternative monomers and heterologously expressed enzymes are shown in bold. PAL, phenylalanine ammonia-lyase; C4H, cinnamate 4-hydroxylase; 4CL, 4-coumarate:CoA ligase; HCT, p-Hydroxycinnamoyl-CoA:quinate/shikimate-p-hydroxycinnamoyltransferase; C3’H, p-coumaroyl quinate/shikimate 3’-hydroxylase; CSE, caffeoyl shikimate esterase; CCoAOMT, caffeoyl-CoA O-methyltransferase; CCR, cinnamoyl-CoA reductase; F5H, ferulate 5-hydroxylase/CAld5H, coniferaldehyde 5-hydroxylase; COMT, koffeinsyre O-methyltransferase; CAD, cinnamylalkoholdehydrogenase; FMT, feruloyl-CoA monolignoltransferase; PMT, p-coumaroyl-CoA monolignoltransferase.
i betragtning af at lignin er en vigtig recalcitrance faktor i træ delignification processer, har store forskningsindsats været afsat til optrævle lignin biosyntetiske vej, og at undersøge virkningerne af forstyrrelser af lignin biosyntesen gener på lignin mængde og sammensætning, og på træforarbejdning effektivitet. Mens ændringer i gener, der spænder fra dem, der koder for transkriptionsfaktorer op til dem, der koder for iltning, har resulteret i ændret ligninindhold, sammensætning eller aflejring (Eriksson et al., 2000; Li Y. H. et al., 2003; Liang et al., 2008; Lu et al., 2013; Lin et al., 2016; Et Al., 2017; Yang et al., 2017; Obudulu et al., 2018), vil denne gennemgang fokusere på resultaterne opnået ved at konstruere lignin biosyntetiske gener.
Engineering af Ligninvejen
tabel 1 giver en oversigt over de forskellige undersøgelser af nedregulerede eller muterede ligninbiosyntetiske gener i poppel, fyr, eukalyptus og birk med de resulterende virkninger på træforarbejdningseffektiviteter, når de bestemmes. Reduktion af aktiviteten af ethvert trin i lignin biosyntetisk vej, startende fra PAL op til CAD, kan resultere i en reduktion i ligninindholdet (tabel 1). Flere parametre påvirker graden af ligninreduktion, såsom målgenet og graden af nedregulering af aktiviteten, hvilket igen afhænger af effektiviteten af den anvendte lyddæmpningskonstruktion, størrelsen på genfamilien og redundans inden for genfamilien. Generelt resulterer nedreguleringen af trinnene fra C4H op til CCR i en mere dramatisk reduktion i ligninmængde (Hu et al., 1999; Meyermans et al., 2000; Jong et al., 2000; Li L. et al., 2003; Jia et al., 2004; Lu et al., 2004; LEPL Krist et al., 2007; Coleman et al., 2008a, b; Bjurhager et al., 2010; Mansfield et al., 2012; Ralph et al., 2012; Min et al., 2014; Van Acker et al., 2014; Jens et al., 2015, 2018; Saleme et al., 2017; Jens et al., 2017) end nedregulering af F5H, COMT og CAD (Van Doorsselaere et al., 1995; Baucher et al., 1996; Lapierre et al., 1999; Jouanin et al., 2000; Van Acker et al., 2014; Jørgen et al., 2018). Ligninreduktion kan være forbundet med en stigning i S/G, såsom i C3 ‘ h- (Coleman et al., 2008a; Ralph et al., 2012) og CCoAOMT-nedregulerede træer (Meyermans et al., 2000) eller et fald i S/G – forhold, såsom i CSE – (Saleme et al., 2017), og COMT-nedregulerede træer (Van Doorsselaere et al., 1995; Lapierre et al., 1999; Jouanin et al., 2000). Interessant nok viste det sig, at lav-lignin 4CL-nedregulerede poplars havde en stigning i S/g (Min et al., 2014; Jens et al., 2017), et fald i S/G (Voelker et al., 2010; Jens et al., 2015) eller forhold, der kan sammenlignes med vildtype (Hu et al., 1999; Li L. et al., 2003). Denne varians kan ikke forbindes med promotoren eller den metode, der anvendes til nedregulering, hvilket antyder, at forskelle i graden af lyddæmpning, vækstbetingelser eller udviklingstilstand påvirker dette træk. På den anden side er de stærkeste virkninger på H/G/S ligninsammensætning blevet observeret for træer nedreguleret i C3 ‘ H og HCT, som deponerer lignin med store stigninger i h-enhedsindhold (Coleman et al., 2008a; Ralph et al., 2012; Vanholme et al., 2013A), mens træer, der overudtrykker F5H, producerer lignin stærkt beriget i S-enheder (Franke et al., 2000; Li L. et al., 2003; Mads et al., 2009), og træer, der er nedreguleret i COMT, har dramatisk reduceret s-enhedens biosyntese (Van Doorsselaere et al., 1995; Lapierre et al., 1999).

tabel 1. Oversigt over skovtræer med modificeret ekspression af ligninbiosyntesegener.
biomassens behandlingseffektivitet kan også modificeres ved øget inkorporering af molekyler, der generelt repræsenterer mindre komponenter i lignin af vildtypeplanter. Inkorporering af ferulinsyre i CCR-mangelfulde træer resulterer i dannelsen af acetalbindinger i ligninpolymeren, som let spaltes i sure biomasseforbehandlinger (LEPL Larsen et al., 2007; Ralph et al., 2008; Van Acker et al., 2014). Faktisk korrelerede niveauerne af ferulinsyre i lignin positivt med en højere saccharificeringseffektivitet (Van Acker et al., 2014). 5-hydroksyksiferylalkohol og 5-hydroksiksiferaldehyd i lignin af COMT-mangelfulde poplars (Van Doorsselaere et al., 1995; Lapierre et al., 1999; Jouanin et al., 2000; Morreel et al., 2004; Lu et al., 2010) giver anledning til bindinger, der potentielt forhindrer kovalente forbindelser mellem lignin og polysaccharidhydroksylgrupperne., 2010; Vanholme et al., 2012; Nishimura et al., 2018). På den anden side resulterer COMT-mangel også i et mere kondenseret lignin på grund af de relativt højere niveauer af de kondenserede karrusel-og karrusel-5-bindinger og de lavere niveauer af karrusel-O-4-bindinger, når s-enhedsfrekvensen falder. Kemisk pulping af træ afledt af poplars stærkt nedreguleret for COMT resulterede i et højere pulpudbytte, opvejet af det resterende ligninindhold i papirmassen. Disse træer havde et lavere lignin og et højere celluloseindhold (Jouanin et al., 2000). På den anden side havde popler, der var beskedent nedreguleret for COMT, et stort fald i pulpudbyttet, formodentlig fordi ligninindholdet var forblevet normalt, mens ligninet havde en højere frekvens af kondenserede bindinger, der negativt påvirkede ligninekstraktionen (Lapierre et al., 1999; Pilatus et al., 2002). Inkorporeringen af cinnamaldehyder i ligninpolymeren i CAD-mangelfulde træer resulterer i kortere ligninpolymerkæder, dermed en højere andel af frie phenolendegrupper, der øger opløseligheden af polymeren i alkali. Inkorporeringen af cinnamaldehyder i ligninpolymeren reducerer formodentlig også den kovalente interaktion mellem den alifatiske kæde og hemicellulose, hvilket igen gør ligninet mere opløseligt. På grund af det udvidede konjugerede system, der genereres, når et cinnamaldehyd-Kurt-o-4 parres med en anden monomer, bliver den aromatiske etherbinding af det inkorporerede cinnamaldehyd mere modtagelig for alkalisk spaltning (Lapierre et al., 1989; Van Acker et al., 2017).
Ligninpolymerisering er en kombinatorisk radikal koblingsproces, der tillader, at en bred vifte af phenolforbindelser naturligt inkorporeres i ligninpolymeren (Boerjan et al., 2003; Vanholme et al., 2019). Forskere har forsøgt at skræddersy ligninmængden og sammensætningen for at forbedre biomasseforarbejdning ved ekspression af heterologe gener, der sigter mod biosyntese og inkorporering af forskellige kompatible phenolforbindelser som alternative monolignoler i ligninpolymeren (Ralph, 2006; Vanholme et al., 2012; Mottiar et al., 2016; Mahon og Mansfield, 2018). Et eksempel er introduktionen af gener, der er nødvendige for S-enhedsbiosyntese i fyr; den samtidige ekspression af F5H, COMT og CAD introducerede med succes s-enheder i Pinus radiata., 2015; Edmunds et al., 2017). Indførelsen af genet, der koder for en monolignol 4-O-methyltransferase (MOMT4) i Poppel fører til dannelsen af 4-o-methylerede coniferyl-og sinapylalkoholer, som ikke kan inkorporeres i den voksende ligninpolymer, fordi de mangler den aromatiske hydroksylgruppe. Dette fører til et stop i ligninpolymerisation og resulterer i træer med lavere ligninindhold og højere saccharificeringseffektivitet (Bhuiya og Liu, 2010; Cai et al., 2016). Poplars er også blevet konstrueret til at indeholde esterbindinger i ligninpolymerens rygrad. Coniferylferulatestere blev introduceret i polymeren via ekspression af et FERULOYL-CoA:MONOLIGNOLTRANSFERASE (FMT) gen afledt af Angelica sinensis (Vilkerson et al., 2014), hvilket fører til en forbedret saccharificeringseffektivitet under forskellige forbehandlingsbetingelser., 2014; Kim et al., 2017; Bhalla et al., 2018), og en forbedret kraftpulpingseffektivitet sammenlignet med vildtype (Jhou et al., 2017). Monolignol p-coumaratestere er også blevet konstrueret i Poppel via ekspression af et rice p-COUMAROYL-CoA:monolignol TRANSFERASE (PMT) gen, hvilket resulterer i en højere frekvens af resistente interunitbindinger og en højere frekvens af G-og S-terminalenheder med frie phenolgrupper (Smith et al., 2015; Sibout et al., 2016). Mens i Arabidopsis resulterede den heterologe ekspression af PMT i en reduceret ligninmængde ledsaget af en øget saccharificeringseffektivitet (Sibout et al., 2016), var der intet fald i ligninmængden i poppel, og saccharificeringseffektiviteten blev ikke bestemt (Smith et al., 2015).
mens flere ændringer af ligninmængden og sammensætningen viste sig at give forbedringer i biomasseforarbejdning, blev disse ændringer ofte ledsaget af en biomasseudbyttestraf (LEPL LARP et al., 2007; Karlslund et al., 2008; Pernille et al., 2009; Voelker et al., 2010; Stout et al., 2014; Van Acker et al., 2014; Sykes et al., 2015; Jens et al., 2018). En nylig metastudie forstyrrede 21 lignin biosyntesegener i P. trichocarpa, og omfattende integreret resultaterne af transcriptomic, proteomic, fluksomic og phenomic data på 221 linjer. Forfatterne konkluderede, at trævækst ikke er forbundet med ligninmængde, underenhedssammensætning eller specifikke forbindelser., 2018), men snarere korreleret med tilstedeværelsen af kollapsede skibe (Coleman et al., 2008a, b; B. et Al., 2009; Voelker et al., 2010; Vargas et al., 2016; De Meester et al., 2018), aktiveringen af en cellevægsintegritetsvej (Bonavit et al., 2014) og/eller akkumulering af kemiske hæmmere (Gallego-Giraldo et al., 2011; Muro-Villanueva et al., 2019).
mens der er gjort en betydelig indsats for at reducere ligninindholdet ved nedregulering af ligninbiosyntetiske gener, undersøgelser af opregulering af ligninvejen og overproduktion af lignin har været knappe. Faktisk viser rapporter om overekspression af F5H et uændret eller endog et fald i ligninindholdet (Huntley et al., 2003; Li L. et al., 2003; Mads et al., 2009; Mansfield et al., 2012; Edmunds et al., 2017). Overekspression af CAD og COMT har resulteret i genhæmning snarere end opregulering, eller der blev ikke påvist nogen effekt på ekspressionsniveauer (Baucher et al., 1996; Lapierre et al., 1999; Jouanin et al., 2000; LEPL Kris et al., 2007; Van Acker et al., 2014). Overekspressionen af r2r3-MYB-transkriptionsfaktorerne PtoMYB92, PtoMYB216 og PtoMYB74 resulterede alle i yderligere lag, tykkere cellevægge samt ektopisk ligninaflejring, og planterne akkumulerede 13-50% mere lignin., 2013; Li C. F. et al., 2015; Li et al., 2018). MYB-overekspressionslinjerne opregulerede konstitutivt ligninbiosyntesevejsgenerne, og mens planter, der overudtrykkede MYB92 og MYB74, havde en biomassestraf, resulterede OVEREKSPRESSIONEN af MYB216 i planter med op til 50% mere lignin og ingen udviklingsfænotype. Da lignin i stigende grad betragtes som en vigtig ressource til bæredygtig produktion af kemikalier (cao et al., 2018) konstruktionen af planter, der overproducerer lignin, bør undersøges nærmere.
feltforsøg
eksemplerne diskuteret ovenfor viser tydeligt, at ligninteknik via ned – eller opregulering af phenylpropanoidvejsgener – eller ekspression af heterologe gener – har potentialet til at øge behandlingseffektiviteten af lignocellulosisk biomasse. Af praktiske og lovgivningsmæssige årsager rapporterer de fleste undersøgelser om data opnået ved analyse af træer dyrket i et drivhus. Imidlertid tager eksperimenter med træer dyrket i et drivhus typisk ikke hensyn til udviklingsprocesser som vækstophør og dvaletilstand. Derudover giver drivhuseksperimenter ikke tilstrækkelig indsigt i interaktionen mellem det konstruerede anlæg og miljøfaktorer såsom jordtype, vind og patogener. Forståelse af disse interaktioner er et vigtigt skridt i oversættelsen af forskningsresultater mod kommercielle applikationer. Faktisk fremhæver det arbejde, der er produceret af undersøgelser, for hvilke der blev givet tilladelse til at etablere feltforsøg, vigtige forskelle i fænotype mellem drivhus – og markdyrkede træer. Tabel 1 opsummerer rapporterne om feltforsøg udført med 4CL, CCoAOMT, CCR, COMT og CAD nedregulerede træer.
bekræftelse af potentialet for modificeret lignocellulosisk biomasse som substrat til applikationer viste flere lignin-konstruerede feltforsøg dyrkede træer forbedringer i træforarbejdning. Poplars nedreguleret for CCoAOMT dyrket i 5 år i et feltforsøg i Beijing (Kina), viste en øget glukose-og ksylosefrigivelse ved saccharificering (Vang et al., 2012). Poplars nedreguleret for CCR og dyrket i et feltforsøg i Frankrig, viste sig at være mere modtagelige for kemisk kraftmasse (LEPL Kurt et al., 2007). To yderligere feltforsøg udført i Frankrig og Belgien med CCR-nedregulerede popler resulterede i op til 160% forbedring i ethanolproduktion i samtidige saccharificerings-og fermenteringsanalyser (SSF); planterne havde dog op til 50% reduktion af biomasse (Van Acker et al., 2014). Feltforsøg med CAD-nedreguleret poppel viste også lovende resultater. Disse træer viste lidt mindre lignin end vildtype og viste sig at være mere modtagelige for kraft-delignificering (Lapierre et al., 1999). Konsekvent viste de samme linjer, der blev dyrket i større feltforsøg i Frankrig og Det Forenede Kongerige, et mildt fald i ligninmængden og en forbedring af kraftmasse, der blev anset for kommercielt relevant, da planterne havde brug for 6% mindre alkali for at opnå en delignificering svarende til den for vilde træer (Pilatus et al., 2002).
modstridende rapporter om både biomasseudbytte og efterfølgende behandlingseffektivitet antyder imidlertid, at disse parametre er stærkt påvirket af miljøfaktorer. Mens et feltforsøg udført i Kina ved hjælp af 4CL nedregulerede poplars fandt, at selv med et fald på 28% i ligninindhold sammenlignet med vildtype, havde træerne omkring 8% øget højde., 2013), i overensstemmelse med drivhusundersøgelser (Hu et al., 1999), fandt andre feltforsøg, at 4CL-nedregulerede poplars havde nedsat biomasse og undertiden endda var dværgede (Voelker et al., 2010; Stout et al., 2014; Marchin et al., 2017). Rapporter afviger også med hensyn til nedstrøms forarbejdningseffektivitet af træ afledt af disse 4CL-nedregulerede feltdyrkede popler. Mens der blev fundet op til 100% stigning i sukkergenvinding for 4cl1-nedregulerede træer (35S-drevet antisense 4CL-konstruktion) dyrket på et Bjergsted i USA., 2017), data opnået fra feltundersøgelser udført i Oregon (USA) fandt, at Pt4CL1 promotor-drevet antisense tavs 4cl1 poplars havde ingen forbedring i saccharificeringseffektivitet sammenlignet med vildtype (Voelker et al., 2010). Ligeledes fandt en langsigtet undersøgelse i Vinling (Kina), at 4CL-nedregulerede popler ikke viste en signifikant forbedring i sukkerudbyttet sammenlignet med vildtype., 2012). I begge sidstnævnte tilfælde viste træerne milde fald i ligninmængde, som ikke oversatte til højere behandlingseffektivitet, potentielt på grund af den højere koncentration af ekstrakter, der kunne interferere med Fermatisk aktivitet (Voelker et al., 2010).
feltforsøg har vist, at miljøfaktorer kan påvirke lignificering og gendanne træk til vildtypeniveauer sammenlignet med de niveauer, der blev opnået, da de samme planter blev dyrket i drivhuset. Mens 4CL-nedregulerede træer havde nedsat ligninindhold, når de blev dyrket under drivhusforhold, har analyse af de samme 4CL antisense poplars, men dyrket i marken, ofte vist, at ligninindholdet blev øget sammenlignet med de drivhusdyrkede træer og undertiden endda gendannet til vildtype niveauer (Stout et al., 2014; Jens et al., 2017). Tilsvarende blev ligninniveauerne meget mindre reduceret i CCR-mangelfulde poplars, da de blev dyrket i marken sammenlignet med da de blev dyrket i drivhuset (Van Acker et al., 2014). I det mindste for de CCR-mangelfulde poplars er det muligt, at det højere lignificeringsniveau for markdyrkede træer skyldes, at træprøverne blev taget om vinteren. Når trævæksten ophører om efteråret, træerne har stadig tid til fuldt ud at lignificere deres cellevægge, når træet går i dvale, sammenlignet med drivhusdyrkede træer, der kontinuerligt udvikler sig. Ligninsammensætning har også vist sig at variere mellem drivhus – og markdyrkede lav-lignin træer. 4CL-nedregulerede poplars dyrket i et felt i North Carolina havde lignin med en lavere S/G end da de samme linjer blev dyrket i drivhuset (Stout et al., 2014).samlet set viser disse resultater, at data fra drivhusdyrkede træer ikke let kan ekstrapoleres til markdyrkede træer, hvilket understøtter behovet for feltforsøg på forskellige steder. Nogle linjer præsenterede en udbyttestraf, der gjorde dem mindre interessante til applikationer, hvilket fremhævede behovet for en bedre forståelse af det molekylære grundlag for udbyttestraffen og udviklingen af strategier til at overvinde dette problem.
Lignin har vist sig at spille en vigtig rolle i patogenresistens (Miedes et al., 2014, 2014), og det spiller en central rolle i at lade anlægget transportere vand. Dette antyder, at ligninændringer kan have indflydelse på planternes stresstolerance. Mens yderligere undersøgelse er nødvendig for fuldt ud at tackle denne mulighed, ændrede nedreguleringen af 4CL, COMT og CAD i Poppel ikke dramatisk fodringsydelsen for bladfodrende planteædere (Tiimonen et al., 2005; Brodeur-Campbell et al., 2006; Hjalten et al., 2013). Effekten af nedreguleringen af COMT og CAD i Poppel på plante-insektinteraktioner er også blevet vurderet på markdyrkede træer, og det blev vist, at de ligninmodificerede træer havde normal forekomst af besøgende og fodring af insekter såvel som normale reaktioner på mikrobielle patogener (Pilatus et al., 2002; Halpin et al., 2007). Disse resultater indikerer, at træer med modificeret lignin ikke nødvendigvis lider mere end vilde planter fra skadedyr og sygdomme. Ikke desto mindre viste profilering af endosfærens bakterielle mikrobiom af træ høstet fra markdyrkede, CCR-nedregulerede popler skift i bakteriesamfundet, formodentlig på grund af den ændrede overflod af bestemte phenolmetabolitter i Ksylem (Beckers et al., 2017).
i betragtning af lignins rolle i funktion og struktur er vandforholdene for nogle få lav lignin-modificerede poplars blevet vurderet. 4CL-nedregulerede poplars viste sig at have reduceret hydraulisk ledningsevne, hvilket potentielt interfererede med plantevækst (Marchin et al., 2017). Hydrauliske spændingseksperimenter med popler nedreguleret til CCR, COMT eller CAD viste, at disse planter havde en lavere modstand mod kavitation, samtidig med at de opretholdt normal hydraulisk ledningsevne og vandtransport., 2012). Disse resultater tyder på, at væksten af lav-lignin mutanter kan være påvirket af vand tilgængelighed. Som for enhver ny hybrid opnået fra klassisk avl, er feltforsøg nødvendige for at evaluere feltpræstation og stresstolerance for ligninkonstruerede træer.
udsigterne for Ligninteknik i skovtræer
udførelsen af ligninkonstruerede planter ser ud til at være stærkt påvirket af miljøforholdene. Det er imidlertid uklart, om forskellene mellem drivhusdyrkede og markdyrkede træer eller mellem træer dyrket på forskellige marksteder skyldes forskellige niveauer af genundertrykkelse eller fra interaktion mellem det konstruerede træk med miljøet. Faktisk er ustabil nedregulering en mangel på genhæmningsteknikker, der er baseret på RNAi. Dette ses ved at observere variation i den røde fænotype, der observeres, når bestemte ligninbiosyntesegener, såsom CAD, COMT eller CCR, nedreguleres. Den røde farve er ofte ikke ensartet i hele farven, men vises snarere i patches, der afspejler variable niveauer af genhæmning (LEPL Larsen et al., 2007; Voelker et al., 2010; Van Acker et al., 2014; figur 2). Derudover kan brugen af genhæmningsmetoder potentielt resultere i samtidig lyddæmpning af nært beslægtede genfamiliemedlemmer – måske i forskellige grader – tilsløre fortolkningerne og camouflere virkningerne af nedregulering af individuelle gener.

figur 2. Patchy gen nedregulering af RNAi. Patchy rød fænotype observeret på stammer af CCR-mangelfulde poplars (højre) dyrket i et feltforsøg i Belgien. Den røde del angiver områder med CCR-nedregulering. Træ fra vilde træer er hvidligt (til venstre).
disse problemer kan nu let overvindes ved brug af CRISPR-baserede genredigeringsteknologier, der muliggør stabile tab af funktionsmutationer (knock-outs) i specifikke målgener, hvilket muliggør dissektion af funktionen af individuelle gener i familier. For eksempel viste målretningen af individuelle 4CL-genfamiliemedlemmer i Poppel, at 4cl1 er relateret til lignificering, mens 4cl2 er involveret i proanthocyanidinproduktion (Jhou et al., 2015). Ud over knock-out alleler giver CRISPR-baseret genredigering også mulighed for at oprette nye alleler, der giver delvis reduktion i aktivitet. Dette åbner mulighed for at finjustere niveauet af restaktivitet og omgå den udbyttestraf, der ofte observeres, når ligninmængden falder under et tærskelniveau. En anden lovende vej til ligninteknik i skovtræer muliggjort gennem CRISPR-baseret genomteknik er samtidig redigering af flere gener (allelstablering) for at optimere effektiviteten af biomassebehandling, som eksemplificeret i Arabidopsis, hvor stabling af transaldolase (tra) og comt mutationer, C4H og comt mutationer eller 4CL og comt mutationer resulterede i additive og synergistiske forbedringer i saccharificeringseffektivitet (de Vries et al., 2018). Ja, en systemtilgang i P. trichocarpa forudsiger, at den samtidige nedregulering af PAL og CCoAOMT, eller PAL, C3 ‘ H og CCOAOMT vil forbedre træegenskaber og frigivelse af sukker væsentligt., 2018).
anvendelsen af CRISPR-baseret genomredigering i træforbedring til papirmasse-og papir-og bioraffinaderiindustrien samt til produktion af platformaromater fra den hydrogenolytiske nedbrydning af lignin vil være mest værdifuld, når denne teknologi strategisk kombineres med andre avlsteknikker (figur 3). Faktisk findes der allerede stor variation i ligninmængde og S/G-sammensætning i naturlige populationer af skovtræer (Studer et al., 2011). I betragtning af at begge træk påvirker glukosefrigivelsen ved saccharificering (Yoo et al., 2018), der udnytter denne genetiske mangfoldighed ved konventionel avl, hjulpet af genom-brede associeringsundersøgelser (Porth et al., 2013; Fahrenkrog et al., 2017; Liu et al., 2018), avl med sjældne defekte alleler (BRDA) (Vanholme et al., 2013A) eller genomisk selektion (Yin et al., 2010; Muchero et al., 2015; et al., 2018; Jens et al., 2018), er en værdifuld strategi for at opnå linjer, der har forbedret træforarbejdningseffektivitet. Når elitetræer er opnået ved disse avlsmetoder, genteknologi og CRISPR-baseret genredigering af specifikke gener er en meget lovende vej til yderligere at forbedre disse elitegenotyper uden at bryde deres genetiske sammensætning og uden at gennemgå lange avlscyklusser. I betragtning af det forestående klimanedbrud har vi ikke mere tid til at tabe ved at vedtage disse nye avlsteknikker i vores løb mod den biobaserede Økonomi.
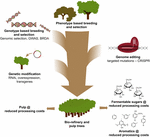
figur 3. Genetisk forbedring af skovtræer gennem en kombination af avlsværktøjer. For at fremskynde den genetiske forbedring af skovtræer til papirmasse-og bioraffinaderiapplikationer skal klassiske og nye avlsværktøjer kombineres smart. Klassisk avl involverer fænotypisk udvælgelse af træer til kontrollerede kryds efterfulgt af fænotypisk udvælgelse. Med fremkomsten af genomsekvensinformation fra mange skovtræer er der udviklet nye strategier såsom genomisk selektion, genom-brede associeringsundersøgelser (GVAR) og avl med sjældne defekte alleler (BRDA) for at fremskynde indfangning og berigelse af DNA-polymorfier forbundet med gavnlige træk. CRISPR-baseret genomredigering gør det muligt at modificere genomet på en måde, der efterligner naturlige polymorfier. Genetisk modifikation involverer stabil integration af fremmed DNA i træet for at overproducere (et) eller nedregulere(a) gen (er). Det er nødvendigt at kombinere de klassiske og nye avlsteknikker for at give tilstrækkeligt træ af høj kvalitet til samfundet.
Forfatterbidrag
alle forfattere, der er anført, har ydet et væsentligt, direkte og Intellektuelt Bidrag til Værket og godkendt det til offentliggørelse.
finansiering
Vi anerkender delvis finansiering fra projektet BIOLEUM (Grant No. 130039) og af SBO-FISH gennem ARBOREF-projektet. AC har modtaget støtte fra FN ‘ s forsknings-og innovationsprogram Horisont 2020 under Marie sk Curie-tilskudsaftalen nr.665501. LdV blev finansieret af Institut for fremme af Innovation gennem videnskab og teknologi i Flandern til et predoktoralt stipendium.
interessekonflikt Erklæring
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
anerkendelser
Vi takker Annick Bleys for at forberede dette manuskript til indsendelse.
Boerjan, Ralph, J. og Baucher, M. (2003). Lignin biosyntese. Annu. Rev. Plant Biol. 54, 519–546.
PubMed abstrakt/Google Scholar
Lapierre, C., Pollet, B., Petit-Conil, M., Toval, G., Romero, J., Pilatus, G., et al. (1999). Strukturelle ændringer af ligniner i transgene poplars med deprimeret cinnamylalkoholdehydrogenase eller koffeinsyre O-methyltransferaseaktivitet har en modsat indvirkning på effektiviteten af industriel kraftmasse. Plant Physiol. 119, 153–163.
PubMed abstrakt / Google Scholar
Ralph, J. (2006). “Hvad gør en god monolignol erstatning?, “i videnskaben og historien om plantecellevæggen – biosyntese, struktur og funktion, Red. T. Hayashi (Boca Raton, FL: Brunvandringspresse).
Google Scholar