tanulási célok
e szakasz végére képes lesz:
- magyarázza el a glikolízis folyamatait
- írja le a piruvát molekula útját a Krebs-cikluson keresztül
- magyarázza az elektronok transzportját az elektrontranszport láncon keresztül
- írja le az ATP-termelés folyamatát oxidatív foszforilációval
- foglalja össze a glükoneogenezis folyamatát
a szénhidrátok szén -, hidrogén-és oxigénatomokból álló szerves molekulák. A szénhidrátok családja mind egyszerű, mind összetett cukrokat tartalmaz. A glükóz és a fruktóz az egyszerű cukrok, a keményítő, a glikogén és a cellulóz pedig a komplex cukrok példái. A komplex cukrokat poliszacharidoknak is nevezik, és több monoszacharid molekulából állnak. A poliszacharidok energiatárolóként (pl. keményítő és glikogén) és szerkezeti összetevőként (pl. kitin a rovarokban és cellulóz a növényekben) szolgálnak.
az emésztés során a szénhidrátokat egyszerű, oldható cukrokra bontják, amelyek a bél falán keresztül szállíthatók a keringési rendszerbe, hogy az egész testben szállíthatók legyenek. A szénhidrát emésztés a szájban kezdődik a nyál-amiláz keményítőkre gyakorolt hatásával, és a vékonybél epitéliumán keresztül felszívódó monoszacharidokkal végződik. Miután az abszorbeált monoszacharidokat a szövetekbe szállítják, megkezdődik a sejtlégzés folyamata (1.ábra). Ez a szakasz először a glikolízisre összpontosít, egy olyan folyamatra, ahol a monoszacharid glükóz oxidálódik, felszabadítva a kötéseiben tárolt energiát az ATP előállításához.
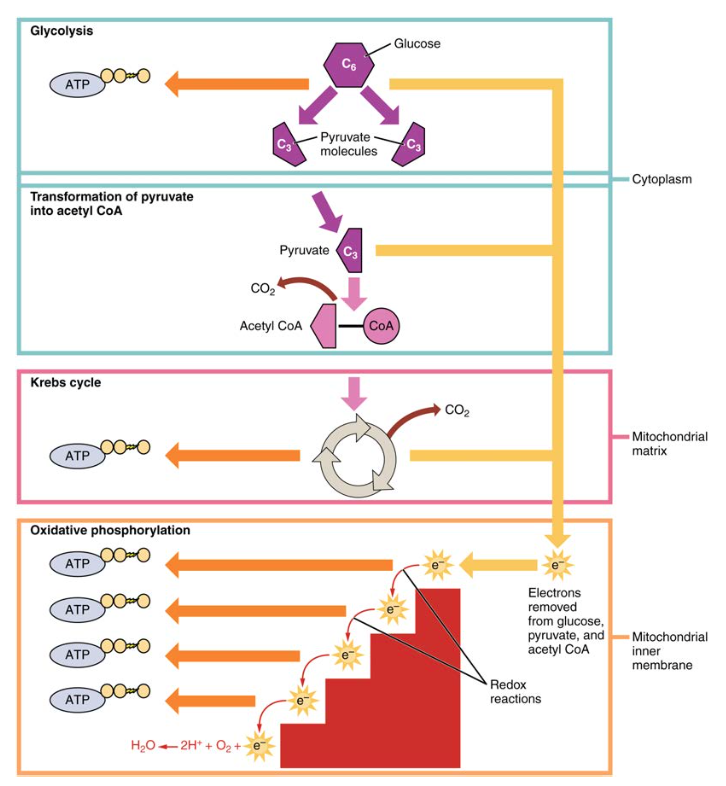
1.ábra. A sejtlégzés glikolízissel, a Krebs-ciklussal és oxidatív foszforilációval oxidálja a glükózmolekulákat, hogy ATP-t termeljen.
glikolízis
a glükóz a szervezet legkönnyebben elérhető energiaforrása. Miután az emésztési folyamatok lebontják a poliszacharidokat monoszacharidokká, beleértve a glükózt is, a monoszacharidok a vékonybél falán keresztül a keringési rendszerbe kerülnek, amely a májba szállítja őket. A májban a hepatociták vagy továbbítják a glükózt a keringési rendszeren keresztül, vagy glikogénként tárolják a felesleges glükózt. A szervezet sejtjei az inzulin hatására felveszik a keringő glükózt, és a glikolízisnek nevezett reakciók sorozatán keresztül a glükózban lévő energia egy részét az ADP-be továbbítják, hogy ATP-t képezzenek (2.ábra). A glikolízis utolsó lépése a piruvát terméket eredményezi.
A glikolízis a glükóz foszforilezésével kezdődik hexokináz glükóz-6-foszfát képződéséhez. Ez a lépés egy ATP-t használ, amely a foszfátcsoport donora. A foszfofruktokináz hatására a glükóz-6-foszfát fruktóz-6-foszfáttá alakul. Ezen a ponton egy második ATP adományozza foszfátcsoportját, fruktóz-1,6-biszfoszfátot képezve. Ez a hat széntartalmú cukor két foszforilált három szénmolekulát, glicerinaldehid-3-foszfátot és dihidroxi-aceton-foszfátot képez, amelyek mindkettő glicerinaldehid-3-foszfáttá alakul. A glicerinaldehid-3-foszfátot a sejtben jelen lévő dihidrogén-foszfát által adományozott csoportokkal tovább foszforiláljuk, hogy az 1,3-biszfoszfoglicerát három szénmolekulát képezzük. Ennek a reakciónak az energiája a glicerinaldehid-3-foszfát oxidációjából (elektronok eltávolításából) származik. A piruváthoz vezető reakciók sorozatában a két foszfátcsoportot ezután két ADP-be viszik át, hogy két ATP-t képezzenek. Így a glikolízis két ATP-t használ, de négy ATP-t generál, két ATP és két piruvát molekula nettó nyereségét eredményezve. Oxigén jelenlétében a piruvát tovább folytatódik a Krebs-ciklushoz (más néven citromsav-ciklushoz vagy trikarbonsav-ciklushoz (TCA), ahol további energiát extrahálnak és továbbítanak.
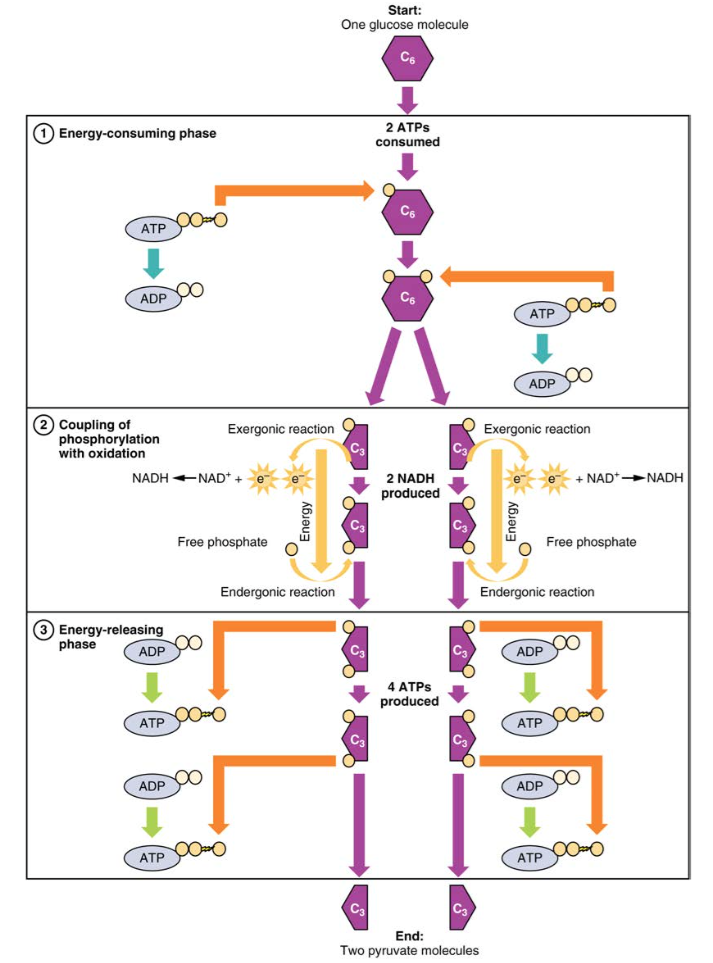
2.ábra. A glikolízis energiaigényes fázisában két ATP-t fogyasztanak, két foszfátot visznek át a glükózmolekulába. A glükózmolekula ezután két három szénvegyületre bomlik, amelyek mindegyike foszfátot tartalmaz. A második fázisban további foszfátot adunk a három szénvegyületekhez. Ennek az endergonikus reakciónak az energiáját két elektron eltávolítása (oxidációja) biztosítja minden három szénvegyületből. Az energia felszabadító fázis során a foszfátokat eltávolítják mind a három szénvegyületből, mind pedig négy ATP molekula előállítására használják.
nézze meg ezt a videót a glikolízisről:
A glikolízis két fázisra osztható: energiafogyasztás (más néven kémiai alapozás) és energiatermelés. Az első fázis az energiafogyasztó fázis, ezért két ATP molekulára van szükség a reakció elindításához minden glükózmolekulára. A reakció vége azonban négy ATP-t eredményez, ami két ATP energiamolekula nettó nyereségét eredményezi.
A glikolízis a következő egyenlettel fejezhető ki:
\text{Glucose}+2\text{ATP}+2\text{NAD}^{+}+4\text{ADP}+2\text{P}_{i}\to\text{Pyruvate}+4\text{ATP}+2\text{NADH}+2\text{H}^{+}
Ez az egyenlet kimondja, hogy a glükóz az ATP—vel (az energiaforrással), a NAD+—val (egy elektron akceptorként szolgáló koenzim) és a szervetlen foszfáttal kombinálva két piruvátmolekulára bomlik, négy ATP-molekulát hozva létre-két ATP-és két energiát tartalmazó NADH-koenzim nettó hozamához. Az ebben a folyamatban előállított NADH-t később felhasználják ATP előállítására a mitokondriumokban. Fontos, hogy ennek a folyamatnak a végére egy glükózmolekula két piruvátmolekulát, két nagy energiájú ATP molekulát és két elektronhordozó NADH molekulát hoz létre.
a glikolízis következő megbeszélései magukban foglalják a reakciókért felelős enzimeket. Amikor a glükóz belép a sejtbe, az enzim hexokináz (vagy glükokináz, a májban) gyorsan hozzáad egy foszfátot, hogy glükóz-6-foszfáttá alakítsa. A kináz egy olyan típusú enzim, amely foszfátmolekulát ad a szubsztráthoz (ebben az esetben glükóz, de más molekulákra is igaz lehet). Ez a konverziós lépés egy ATP-t igényel, és lényegében csapdába ejti a sejtben lévő glükózt, megakadályozva annak visszajutását a plazmamembránon keresztül, ezáltal lehetővé téve a glikolízis folytatását. Ezenkívül a vérben magasabb glükózszinttel rendelkező koncentrációs gradiens fenntartására is szolgál, mint a szövetekben. Ennek a koncentrációs gradiensnek a megállapításával a vér glükózja nagy koncentrációjú területről (a vér) alacsony koncentrációjú területre (a szövetekre) áramolhat, amelyet fel kell használni vagy tárolni. A hexokináz a test szinte minden szövetében megtalálható. A glükokináz viszont olyan szövetekben expresszálódik, amelyek aktívak, ha magas a vércukorszint, például a májban. A hexokináz nagyobb affinitással rendelkezik a glükózhoz, mint a glükokináz, ezért gyorsabban képes átalakítani a glükózt, mint a glükokináz. Ez akkor fontos, ha a glükóz szintje nagyon alacsony a szervezetben, mivel lehetővé teszi a glükóz számára, hogy előnyben részesítse azokat a szöveteket, amelyek többet igényelnek.
a glikolízis első fázisának következő lépésében a glükóz-6-foszfát izomeráz enzim a glükóz-6-foszfátot fruktóz-6-foszfáttá alakítja. A glükózhoz hasonlóan a fruktóz is hat széntartalmú cukor. A foszfofruktokináz-1 enzim ezután még egy foszfátot ad hozzá, hogy a fruktóz-6-foszfátot fruktózvá alakítsa-1-6-biszfoszfát, egy másik hat széntartalmú cukor, egy másik ATP molekula felhasználásával. Az aldoláz ezután lebontja ezt a fruktózt-1-6-biszfoszfát két három szénmolekulába, glicerinaldehid-3-foszfátba és dihidroxi-aceton-foszfátba. A trioszefoszfát izomeráz enzim ezután átalakítja a dihidroxi-aceton-foszfátot egy második gliceraldehid-3-foszfát molekulává. Ezért ennek a kémiai alapozó vagy energiaigényes fázisnak a végére egy glükózmolekulát két gliceraldehid – 3-foszfát molekulára bontunk.
a glikolízis második fázisa, az energiatermelő fázis létrehozza azt az energiát, amely a glikolízis terméke. A gliceraldehid-3-foszfát-dehidrogenáz a
energiaigényes fázis során keletkező három szén-dioxid-gliceraldehid-3-foszfátot 1,3-biszfoszfogliceráttá alakítja. Ez a reakció felszabadít egy elektront, amelyet a NAD+ felvesz, hogy NADH molekulát hozzon létre. A NADH egy nagy energiájú molekula, mint az ATP, de az ATP-vel ellentétben a sejt nem használja energia pénznemként. Mivel két gliceraldehid-3-foszfát molekula van, két NADH molekulát szintetizálunk ebben a lépésben. Ezt követően minden 1,3-biszfoszfoglicerátot defoszforilálunk (azaz foszfátot távolítunk el) a foszfoglicerát-kinázzal 3-foszfogliceráttá. Az ebben a reakcióban felszabaduló minden foszfát képes átalakítani az ADP egy molekuláját egy nagy energiájú ATP molekulává, ami két ATP molekula nyereségét eredményezi.
a foszfoglicerát mutáz enzim ezután a 3-foszfoglicerát molekulákat 2-foszfogliceráttá alakítja. Az enoláz enzim ezután a 2-foszfoglicerát molekulákra hat, hogy foszfoenolpiruvát molekulákká alakítsa őket. A glikolízis utolsó lépése a két foszfoenolpiruvát molekula defoszforilálása piruvát kinázzal, hogy két piruvátmolekulát és két ATP molekulát hozzon létre.
összefoglalva, egy glükózmolekula két piruvátmolekulára bomlik, és glikolízissel két nettó ATP-molekulát és két NADH-molekulát hoz létre. Ezért a glikolízis energiát generál a sejt számára, és olyan piruvátmolekulákat hoz létre, amelyek tovább feldolgozhatók az aerob Krebs-cikluson keresztül (más néven citromsav-ciklus vagy trikarbonsav-ciklus); fermentációval tejsavvá vagy alkohollá alakul (élesztőben); vagy később glükóz szintézisére használják glükoneogenezis útján.
anaerob légzés
Ha az oxigén korlátozott vagy hiányzik, a piruvát anaerob útvonalba lép. Ezekben a reakciókban a piruvát tejsavvá alakítható. Amellett, hogy további ATP-t generál, ez az út arra szolgál, hogy a piruvát koncentrációját alacsonyan tartsa, így a glikolízis folytatódik, és oxidálja a NADH-t a glikolízishez szükséges NAD+ – ba. Ebben a reakcióban a tejsav az oxigént helyettesíti végső elektron akceptorként. Az anaerob légzés a test legtöbb sejtjében akkor fordul elő, ha az oxigén korlátozott, vagy a mitokondriumok hiányoznak vagy nem működnek. Például, mivel az eritrocitáknak (vörösvértesteknek) nincs mitokondriumuk, anaerob légzésből kell előállítaniuk ATP-jüket. Ez az ATP-termelés hatékony útja rövid ideig, másodpercektől néhány percig. A termelt tejsav diffundál a plazmába, és a májba kerül, ahol a Cori-cikluson keresztül piruváttá vagy glükózzá alakul. Hasonlóképpen, amikor egy személy gyakorol, az izmok gyorsabban használják az ATP-t, mint az oxigén. A gyors ATP-termelés a glikolízistől és a tejsavtermeléstől függ.
aerob légzés
oxigén jelenlétében a piruvát beléphet a Krebs-ciklusba, ahol az elektronok a piruvátból a NAD+, GDP és FAD receptorokba kerülnek, a szén-dioxid pedig “hulladéktermék” (3.ábra). A NADH és a FADH2 továbbadja az elektronokat az elektrontranszport Láncnak, amely az átadott energiát ATP előállítására használja fel. Az elektrontranszport lánc terminális lépéseként az oxigén a terminális elektron akceptor, és vizet hoz létre a mitokondriumokban.
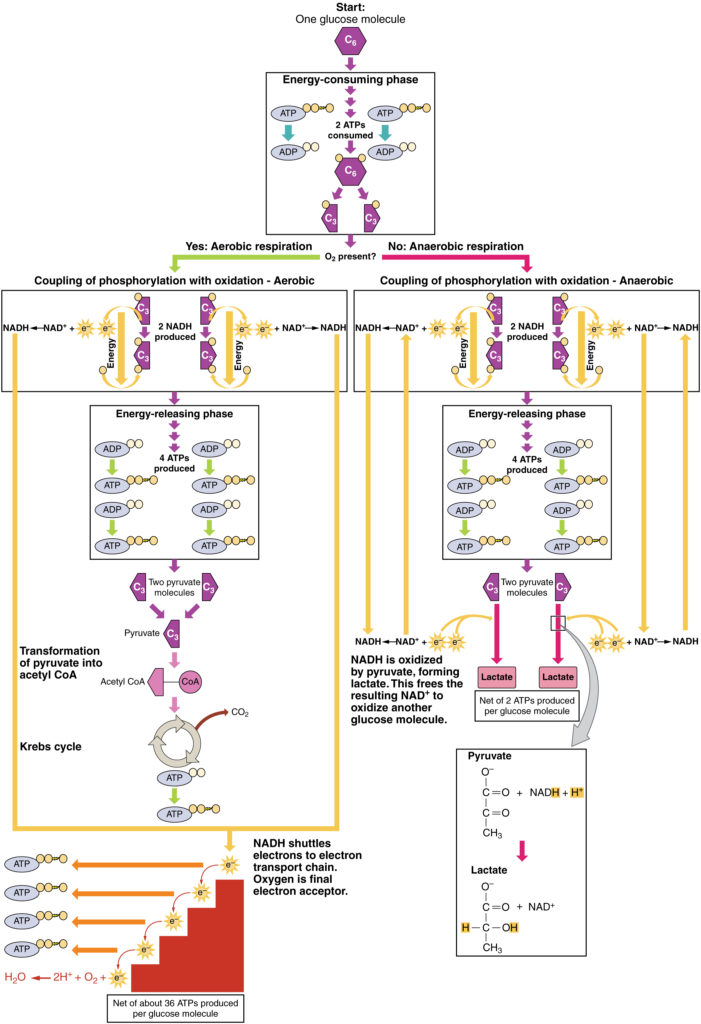
3. ábra. Kattintson a nagyobb kép megtekintéséhez. Az anaerob légzés folyamata a glükózt két laktátmolekulává alakítja oxigén hiányában vagy a mitokondriumok nélküli eritrocitákon belül. Az aerob légzés során a glükóz két piruvátmolekulává oxidálódik.
Krebs-ciklus / citromsav-ciklus / trikarbonsav-ciklus
a glikolízis során keletkező piruvátmolekulák a mitokondriális membránon keresztül a belső mitokondriális mátrixba kerülnek, ahol enzimek metabolizálják őket a Krebs-ciklusnak nevezett úton (4.ábra). A Krebs-ciklust általában citromsav-ciklusnak vagy trikarbonsav-ciklusnak (TCA) is nevezik. A Krebs-ciklus során nagy energiájú molekulák, köztük ATP, NADH és FADH2 jönnek létre. A NADH és a FADH2 ezután elektronokat vezet át a mitokondriumokban lévő elektrontranszport láncon, hogy több ATP molekulát hozzon létre.
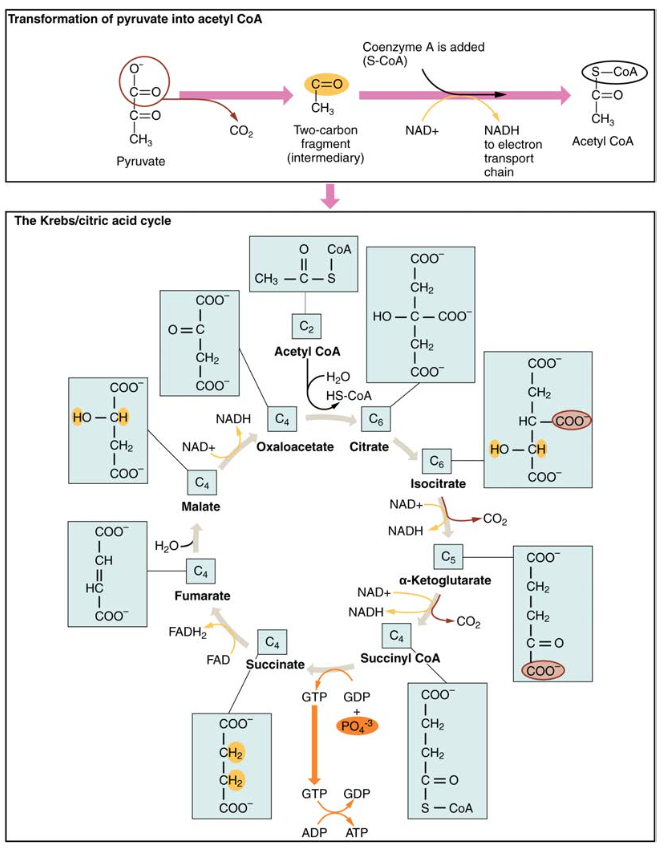
4.ábra. A Krebs-ciklus során minden glikolízis által generált piruvát két szénatomos acetil-CoA molekulává alakul. Az acetil-CoA szisztematikusan feldolgozódik a ciklus során, és nagy energiájú NADH, FADH2 és ATP molekulákat termel.
nézze meg ezt az animációt, hogy megfigyelje a Krebs ciklust.
a glikolízis során keletkező három szén-dioxid-piruvát molekula a citoplazmából a mitokondriális mátrixba mozog, ahol a piruvát-dehidrogenáz enzim két szén-acetil-koenzim A (acetil-CoA) molekulává alakítja. Ez a reakció oxidatív dekarboxilezési reakció. A három szén-dioxid-piruvátot két szén-dioxid-acetil-CoA molekulává alakítja, szén-dioxidot szabadít fel, és két elektront továbbít, amelyek nad+ – val kombinálva NADH-t képeznek. Acetil-CoA belép a Krebs-ciklus kombinálva egy négy szén-dioxid-molekula, oxaloacetát, hogy a hat szén-dioxid-molekula citrát, vagy citromsav, ugyanakkor felszabadító koenzim egy molekula.
a hat szén-citrát molekulát szisztematikusan öt szén-dioxid-molekulává, majd négy szén-dioxid-molekulává alakítják, oxaloacetáttal, a ciklus kezdetével. Az út során minden citrát molekula egy ATP-t, egy FADH2-t és három NADH-t termel. A FADH2 és a NADH belép a belső mitokondriális membránban található oxidatív foszforilációs rendszerbe. Ezenkívül a Krebs ciklus biztosítja a kiindulási anyagokat a fehérjék és zsírok feldolgozásához és lebontásához.
a Krebs-ciklus elindításához a citrát-szintáz kombinálja az acetil-CoA-t és az oxaloacetátot, hogy hat szén-dioxid-citrát molekulát képezzen; a CoA ezután felszabadul, és egy másik piruvát-molekulával kombinálhatja a ciklust. Az akonitáz enzim a citrátot izocitráttá alakítja. Az oxidatív dekarboxilezés két egymást követő lépésében két CO2-és két NADH-molekula keletkezik, amikor az izocitrát-dehidrogenáz az izocitrátot öt szén-dioxid-ketoglutaráttá alakítja, amelyet azután katalizálnak és négy szén-dioxid-szukcinil-CoA-vá alakítanak át a CA-ketoglutarát-dehidrogenáz által. A szukcinil-CoA dehidrogenáz enzim ezután a szukcinil-CoA-t szukcináttá alakítja, és nagy energiájú GTP molekulát képez, amely energiáját az ADP-hez továbbítja ATP előállításához. A szukcinát-dehidrogenáz ezután a szukcinátot fumaráttá alakítja, fadh molekulát képezve2. A fumaráz ezután a fumarátot maláttá alakítja, amely a malát-dehidrogenáz ezután oxaloacetáttá alakul, miközben a NAD+ – ot NADH-ra redukálja. Az oxaloacetát ezután készen áll a következő acetil-CoA-val való kombinálásra a Krebs-ciklus újbóli elindításához (lásd a 4.ábrát). A ciklus minden fordulójára három NADH, egy ATP (GTP-n keresztül) és egy FADH2 jön létre. A piruvát minden szénje Co-vá alakul2, amely az oxidatív (aerob) légzés melléktermékeként szabadul fel.
oxidatív foszforiláció és az elektrontranszport lánc
az elektrontranszportlánc (ETC) a Krebs-ciklus által termelt NADH-t és FADH-t használja az ATP előállításához. A NADH és a FADH2 elektronjai a belső mitokondriális membránba ágyazott fehérjekomplexeken keresztül enzimatikus reakciók sorozatával kerülnek átvitelre. Az elektrontranszport lánc négy enzimkomplexből (komplex I – komplex IV) és két koenzimből (ubiquinon és citokróm c) áll, amelyek elektronhordozóként és protonpumpaként működnek, amelyeket a H+ ionok átvitelére használnak a belső és külső mitokondriális membránok közötti térbe (5.ábra). Az ETC összekapcsolja az elektronok átvitelét egy donor (például NADH) és egy elektron akceptor (például O2) között a protonok (H+ ionok) átvitelével a belső mitokondriális membránon keresztül, lehetővé téve az oxidatív foszforiláció folyamatát. Oxigén jelenlétében az energiát fokozatosan vezetik át az elektronhordozókon, hogy fokozatosan összegyűjtsék a foszfát ADP-hez való csatlakoztatásához és az ATP előállításához szükséges energiát. A molekuláris oxigén szerepe, O2, mint az etc terminális elektron akceptora. Ez azt jelenti, hogy miután az elektronok áthaladtak az egész ETC-n, át kell adni őket egy másik, különálló molekulának. Ezek az elektronok, az O2 és a H+ ionok a mátrixból új vízmolekulákat alkotnak. Ez az alapja annak, hogy oxigént kell lélegeznie. Oxigén nélkül az elektron áramlása az ETC-n keresztül megszűnik.
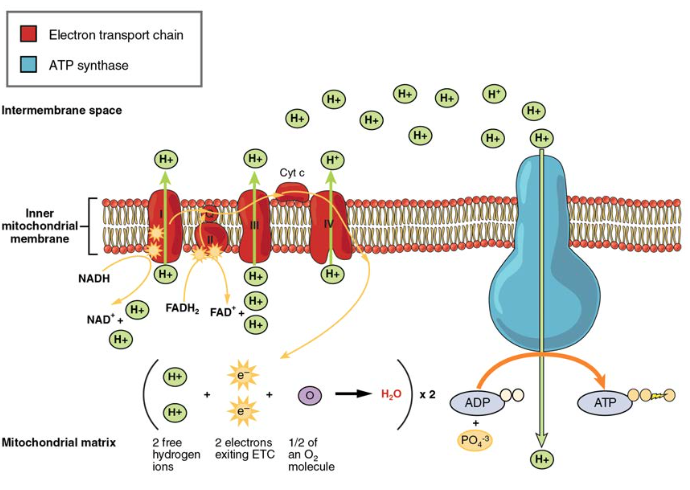
5.ábra. Az elektrontranszport lánc olyan elektronhordozók és ionszivattyúk sorozata, amelyeket H+ ionok szivattyúzására használnak a belső mitokondriális mátrixból.
nézze meg ezt a videót, hogy megismerje az elektrontranszport láncot.
a NADH-ból és a FADH-ból felszabaduló elektronokat az egyes hordozók a lánc mentén vezetik át, amelyek az elektron befogadásakor redukálódnak, és oxidálódnak, amikor továbbadják a következő hordozónak. Ezen reakciók mindegyike kis mennyiségű
energiát szabadít fel, amelyet H+ ionok pumpálására használnak a belső membránon. Ezeknek a protonoknak a felhalmozódása a membránok közötti térben protongradienst hoz létre a mitokondriális mátrixhoz képest.
a belső mitokondriális membránba ágyazva egy csodálatos protein pórus komplex, az úgynevezett ATP szintáz. Valójában ez egy turbina, amelyet a H+ ionok áramlása táplál a belső membránon keresztül egy gradiensen keresztül a mitokondriális mátrixba. Amint a H + ionok áthaladnak a komplexen, a komplex tengelye forog. Ez a rotáció lehetővé teszi az ATP-szintáz más részeinek az ADP és a Pi ATP létrehozására való ösztönzését. Az aerob légzés során glükózmolekulánként előállított ATP teljes számának elszámolásakor fontos megjegyezni a következő pontokat:
- két ATP nettó termelődik glikolízis útján (négy termelődik, kettő pedig az energiafogyasztási szakaszban fogyasztódik). Ezt a két ATP-t azonban arra használják, hogy a glikolízis során keletkező NADH-t a citoplazmából a mitokondriumokba szállítsák. Ezért az ATP nettó termelése a glikolízis során nulla.
- a glikolízis után minden fázisban az ATP, NADH és FADH2 mennyiségét meg kell szorozni kettővel, hogy tükrözze, hogy az egyes glükózmolekulák hogyan termelnek két piruvátmolekulát.
- az ETC-ben körülbelül három ATP keletkezik minden oxidált NADH-ra. Minden oxidált FADH-hoz azonban csak körülbelül két ATP-t állítanak elő2. A FADH2 elektronjai kevesebb ATP-t termelnek, mert az ETC (II.komplex) alacsonyabb pontján kezdődnek, mint a NADH (I. komplex) elektronjai (5. ábra).
ezért minden aerob légzésbe lépő glükózmolekula nettó 36 ATP-t termel (Lásd 6.ábra).
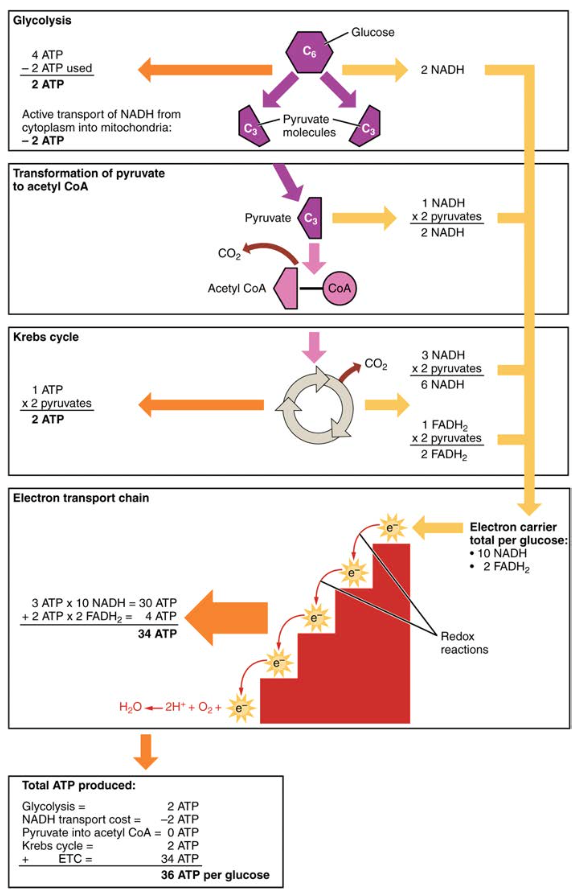
6.ábra. A szénhidrát anyagcsere magában foglalja a glikolízist, a Krebs-ciklust és az elektrontranszport láncot.
glükoneogenezis
a glükoneogenezis a piruvátból, laktátból, glicerinből vagy az alanin vagy glutamin aminosavakból származó új glükózmolekulák szintézise. Ez a folyamat elsősorban a májban zajlik alacsony glükózszint alatt, azaz éhezés, éhezés, alacsony szénhidráttartalmú étrend mellett. Tehát felmerül a kérdés, hogy a test miért hozna létre valamit, amit éppen elég sok erőfeszítést tett a lebontására? Bizonyos kulcsfontosságú szervek, beleértve az agyat is, csak glükózt használhatnak energiaforrásként; ezért elengedhetetlen, hogy a test minimális vércukorszint-koncentrációt tartson fenn. Amikor a vércukorszint az adott pont alá esik, a máj új glükózt szintetizál, hogy a vérkoncentrációt normálisra emelje.
a glükoneogenezis nem egyszerűen a glikolízis fordítottja. Van néhány fontos különbség (7. ábra). A piruvát a glükoneogenezis gyakori kiindulási anyaga. Először a piruvátot oxaloacetáttá alakítják. Az oxaloacetát ezután szubsztrátként szolgál a foszfoenolpiruvát-karboxikináz (PEPCK) enzim számára, amely az oxaloacetátot foszfoenolpiruváttá (PEP) alakítja át. Ebből a lépésből a glükoneogenezis majdnem a glikolízis fordítottja. A PEP visszaalakul 2-foszfogliceráttá, amely 3-foszfogliceráttá alakul. Ezután a 3-foszfoglicerátot 1,3 biszfoszfogliceráttá, majd gliceraldehid-3-foszfáttá alakítják. A gliceraldehid-3-foszfát két molekulája ezután fruktózt képez-1-6-biszfoszfát, amelyet fruktóz 6-foszfáttá, majd glükóz-6-foszfáttá alakítanak át. Végül egy sor reakció maga termel glükózt. A glükoneogenezisben (a glikolízishez képest) a hexokináz enzimet glükóz-6-foszfatáz, a foszfofruktokináz-1 enzimet pedig fruktóz-1,6-biszfoszfatáz váltja fel. Ez segíti a sejtet a glikolízis és a glükoneogenezis szabályozásában egymástól függetlenül.
amint azt a lipolízis részeként tárgyaljuk, a zsírok glicerinre bonthatók, amelyek foszforilezhetők dihidroxi-aceton-foszfát vagy DHAP formájában. A DHAP vagy beléphet a glikolitikus útba, vagy a máj felhasználhatja a glükoneogenezis szubsztrátjaként.
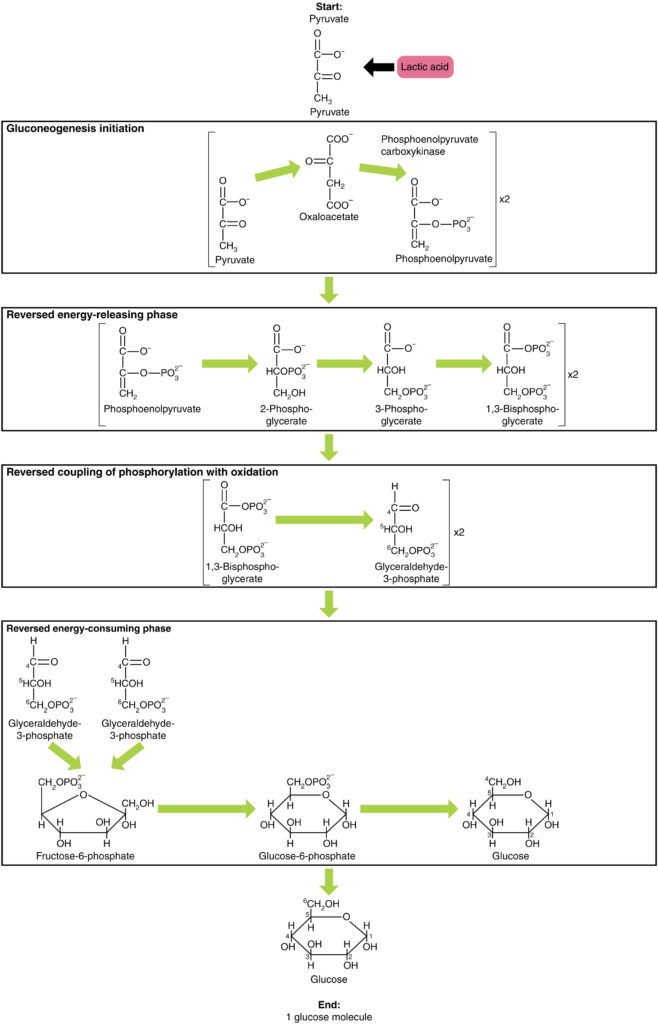
7. ábra. Kattintson a nagyobb kép megtekintéséhez. A glükoneogenezis a piruvátból, laktátból, glicerinből, alaninból vagy glutamátból származó glükóz szintézise.
öregedés és a test anyagcseréje
az emberi test anyagcseréje évtizedenként közel 2 százalékkal csökken 30 éves kor után. A testösszetétel változásai, beleértve a csökkent sovány izomtömeget, leginkább felelősek ennek a csökkenésnek. Az izomtömeg legdrámaibb csökkenése és az anyagcsere sebességének csökkenése 50 és 70 év között következik be. Az izomtömeg elvesztése megegyezik a csökkent erővel, amely hajlamos gátolni az időseket abban, hogy elegendő fizikai aktivitást folytassanak. Ez pozitív visszacsatolási rendszert eredményez, ahol a csökkent fizikai aktivitás még nagyobb izomvesztéshez vezet, tovább csökkentve az anyagcserét.
számos dolog van, amit meg lehet tenni, hogy segítsünk megelőzni az anyagcsere általános hanyatlását, és harcoljunk a csökkenés ciklikus jellege ellen. Ezek közé tartozik a reggeli fogyasztása, a kis étkezések gyakori fogyasztása, sok sovány fehérje fogyasztása, ivóvíz, hogy hidratált maradjon, testmozgás (beleértve az erőnléti edzést) és elegendő alvás. Ezek az intézkedések segíthetnek abban, hogy az energiaszint csökkenjen, és megfékezzék a fokozott kalóriafogyasztás iránti vágyat a túlzott nassolás miatt. Bár ezek a stratégiák nem garantálják az anyagcsere fenntartását, segítenek megelőzni az izomvesztést és növelhetik az energiaszintet. Egyes szakértők azt is javasolják, hogy kerüljék a cukrot, ami túlzott zsírtároláshoz vezethet. A fűszeres ételek és a zöld tea szintén hasznos lehet. Mivel a stressz aktiválja a kortizol felszabadulását, a kortizol pedig lelassítja az anyagcserét, a stressz elkerülése vagy legalábbis a relaxációs technikák gyakorlása szintén segíthet.
fejezet áttekintése
a metabolikus enzimek katalizálják a katabolikus reakciókat, amelyek lebontják az élelmiszerekben található szénhidrátokat. A felszabaduló energiát a testedet alkotó sejtek és rendszerek táplálására használják. A felesleges vagy fel nem használt energiát zsír vagy glikogén formájában tárolják későbbi felhasználás céljából. A szénhidrát-anyagcsere a szájban kezdődik, ahol a nyál-amiláz enzim elkezdi a komplex cukrokat monoszacharidokká bontani. Ezeket ezután a bélmembránon keresztül a véráramba, majd a test szöveteibe lehet szállítani. A sejtekben a glükózt, egy hat széntartalmú cukrot reakciósorozaton keresztül kisebb cukrokká dolgozzák fel, és a molekulában tárolt energia felszabadul. A szénhidrát katabolizmus első lépése a glikolízis, amely piruvátot, NADH-t és ATP-t termel. Anaerob körülmények között a piruvát laktáttá alakítható, hogy a glikolízis működjön. Aerob körülmények között a piruvát belép a Krebs-ciklusba, más néven citromsav-ciklusnak vagy trikarbonsav-ciklusnak. Az ATP mellett a Krebs-ciklus nagy energiájú FADH2 és NADH molekulákat termel, amelyek elektronokat biztosítanak az oxidatív foszforilációs folyamathoz, amely több nagy energiájú ATP molekulát generál. A glikolízis során feldolgozott glükóz minden molekulájára 36 ATPs nettó lehet létrehozni aerob légzéssel.
anaerob körülmények között az ATP-termelés a glikolízis által generáltakra korlátozódik. Míg a glikolízis során összesen négy ATP-t állítanak elő, kettőre van szükség a glikolízis megkezdéséhez, tehát két ATP-molekula nettó hozama van.
alacsony glükózszint esetén, például éhezés, éhezés vagy alacsony szénhidráttartalmú étrend esetén a glükóz szintetizálható laktátból, piruvátból, glicerinből, alaninból vagy glutamátból. Ez a folyamat, az úgynevezett glükoneogenezis, majdnem a glikolízis fordítottja, és glükózmolekulák létrehozására szolgál a glükózfüggő szervek, például az agy számára, amikor a glükózszint a normál alá csökken.
önellenőrzés
válaszoljon az alábbi kérdés(ek) re, hogy lássa, mennyire érti jól az előző szakaszban tárgyalt témákat.
kritikus gondolkodási kérdések
- magyarázza el, hogyan metabolizálódik a glükóz ATP-hoz.
- beszélje meg a sejtek azon mechanizmusát, amelyet koncentrációs gradiens létrehozására alkalmaznak, hogy biztosítsák a glükóz folyamatos felvételét a véráramból.
szószedet
poliszacharidok: sok monoszacharidból álló komplex szénhidrátok
monoszacharid: legkisebb, monomer cukormolekula
nyálamiláz: emésztő enzim, amely a nyálban található, és megkezdi a szénhidrátok emésztését a szájban
sejtlégzés: ATP termelése glükóz oxidációból glikolízissel, a Krebs-ciklus, és oxidatív foszforiláció
glikolízis: metabolikus reakciók sorozata, amely a glükózt piruvátra bontja, és ATP-t termel
piruvát: a glikolízis három széntartalmú végterméke és a kiindulási anyag, amely acetil-CoA-vá alakul, amely belép a
Krebs-ciklusba: más néven a citromsav ciklus vagy a trikarbonsav ciklus, átalakítja piruvát CO2 és nagy energiájú FADH2, NADH, és ATP molekulák
citromsav ciklus vagy trikarbonsav ciklus (TCA): más néven a Krebs ciklus vagy a trikarbonsav ciklus; átalakítja piruvát CO2 és nagy energiájú FADH2, NADH, és ATP molekulák
energia-fogyasztó fázis, első fázisa glikolízis, amelyben két molekula ATP szükséges ahhoz, hogy a piruvát indítsa el a reakciót
glükóz-6-foszfát: a glikolízis első lépésében előállított foszforilált glükóz
hexokináz: celluláris enzim, amely a legtöbb szövetben megtalálható, amely a glükózt glükóz-6-foszfáttá alakítja a sejtbe történő felvételkor
glükokináz: a májban található cellularenzim, amely a glükózt glükóz-6-foszfáttá alakítja a sejtbe történő felvételkor
energiatermelő fázis: a glikolízis második fázisa, amelynek során energiát állítanak elő
terminális elektron akceptor: ATP termelési út, amelyben az elektronok oxidációs-redukciós reakciók sorozatán haladnak át, amelyek vizet képeznek és protongradienst termelnek
Krebs ciklus: más néven a citromsav-ciklus vagy a trikarbonsav-ciklus, átalakítja piruvát CO2 és nagy energiájú FADH2, NADH, és ATP molekulák
elektrontranszport lánc (ETC): ATP termelési útvonal, amelyben az elektronok áthaladnak egy sor oxidációs-redukciós reakciók, hogy vizet képez, és termel egy proton gradiens
oxidatív foszforiláció: folyamat, amely átalakítja a nagy energiájú NADH és FADH2 ATP
ATP szintáz fehérje: pórus komplex, amely létrehozza ATP
glükoneogenezis: a piruvátból vagy más molekulákból származó glükózszintézis folyamata