Bevezetés
a fosszilis erőforrások az energia és a szerves vegyületek fő alapanyagai, és használatuk az éghajlatváltozáshoz kapcsolódó üvegházhatású gázok kibocsátását eredményezi. Az elkövetkező éghajlati összeomlás sürgős átmenetet igényel a fosszilis alapú gazdaságból a bioalapú gazdaságba, amelyben az olaj helyett a lignocellulóz biomasszát használják tüzelőanyagok, vegyi anyagok és anyagok előállítására. A fa a lignocellulóz biomassza fontos forrása; főleg cellulózban, hemicellulózokban és ligninben gazdag másodlagos megvastagodott sejtfalakból áll. Mindhárom polimer értékelhető a bioalapú gazdaságban. A cellulóz a cellulóz-és papíripar forrása, és mind a cellulóz, mind a hemicellulózok depolimerizálhatók monoszacharidjaikba, például bio-etanolba, tejsavba és mosószerekbe (Vanholme et al., 2013b). Mivel a lignin negatívan befolyásolja a fafeldolgozás hatékonyságát ezen alkalmazások felé, a fákat úgy lehet megtervezni, hogy kevesebb lignint halmozzanak fel, hogy jobban alkalmasak legyenek papír és erjeszthető cukrok előállítására. Másrészt a lignint egyre inkább értékes összetevőnek tekintik a bioalapú gazdaságban. Tekintettel arra, hogy a lignin a legnagyobb megújuló aromás forrás a Földön, a biofinomító gazdasági életképessége jelentősen növelhető, ha a lignint is értékelik, és vegyi anyagok előállításához használják fel (Holladay et al., 2007; Tuck et al., 2012; Davis et al., 2013; Ragauskas et al., 2014; Li C. et al., 2015; Van Den Bosch et al., 2015; Rinaldi et al., 2016; Upton és Kasko, 2016; Schutyser et al., 2018).
a lignin polimer monolignolokból áll, amelyeket a fenilpropanoid és a monolignol bioszintetikus úton állítanak elő, a fenilalanin dezaminálásával kezdődő enzimatikus reakciók sorozatával (1.ábra). A monolignolokat a citoplazmában szintetizálják, és az apoplasztba transzlokálják, ahol laccases és peroxidázok hatására monolignol gyökökké dehidrogénezik őket (Berthet et al., 2011; Zhao et al., 2013). Ezek a monolignol-gyökök kombinatorikus módon párosulnak egymással, különféle kémiai kötéseket generálva, mint például az aril-éter kötés (antioxidánsok-o-4), a rezinol kötés (inhibitorok-antioxidánsok) és a fenil-kumarán kötés (antioxidánsok-5) (Boerjan et al., 2003; Ralph et al., 2004; Vanholme et al., 2010). A leggyakoribb monolignolok a hidroxi-cinnamil-alkoholok, a P-kumaril, a tűlevelű és a szinapil-alkoholok, amelyek a lignin polimerbe való beépülésük után generálják a H, G és S egységeket (Bonawitz and Chapple, 2010; Ralph et al., 2019; Vanholme et al., 2019). A lignin építőelemek relatív hozzájárulása a taxonok, a fejlődési stádium, a szövet és a sejttípus, sőt a sejtfal réteg között is változik; a lágyfákból származó lignin (gymnosperms) szinte teljes egészében G egységekből áll, a H egységek kisebb hányadával, míg a keményfákból származó lignin (zárvatermők) a G egységek mellett S egységekkel és H egységek nyomaival rendelkezik (Boerjan et al., 2003; Vanholme et al., 2010, 2019). Ezen hagyományos monolignolok mellett számos más p-hidroxilált aromás molekula is beépíthető a lignin polimerbe különböző szinteken (Vanholme et al., 2019).
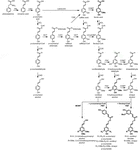
Figure 1. Lignin biosynthetic pathway. Alternative monomers and heterologously expressed enzymes are shown in bold. PAL, phenylalanine ammonia-lyase; C4H, cinnamate 4-hydroxylase; 4CL, 4-coumarate:CoA ligase; HCT, p-Hydroxycinnamoyl-CoA:quinate/shikimate-p-hydroxycinnamoyltransferase; C3’H, p-coumaroyl quinate/shikimate 3’-hydroxylase; CSE, caffeoyl shikimate esterase; CCoAOMT, caffeoyl-CoA O-methyltransferase; CCR, cinnamoyl-CoA reductase; F5H, ferulate 5-hydroxylase/CAld5H, coniferaldehyde 5-hydroxylase; COMT, koffeinsav O-metil-transzferáz; CAD, cinnamil-alkohol-dehidrogenáz; FMT, feruloil-CoA monolignol transzferáz; PMT, p-kumaroil-CoA monolignol transzferáz.
tekintettel arra, hogy a lignin a lignin bioszintetikus útjának feltárására és a lignin bioszintézis gének zavarainak a lignin mennyiségére és összetételére, valamint a fafeldolgozás hatékonyságára gyakorolt hatásainak tanulmányozására nagy erőfeszítéseket tettek. Míg a transzkripciós faktorokat kódolóktól az oxidatív enzimeket kódolókig terjedő gének módosításai megváltozott lignintartalmat, összetételt vagy lerakódást eredményeztek (Eriksson et al., 2000; Li Y. H. et al., 2003; Liang et al., 2008; Lu et al., 2013; Lin et al., 2016; Xu et al., 2017; Yang et al., 2017; Obudulu et al., 2018), Ez a felülvizsgálat a lignin bioszintetikus gének tervezésével kapott eredményekre összpontosít.
A Lignin Útvonal tervezése
az 1.táblázat áttekintést nyújt a nyár, a fenyő, az eukaliptusz és a nyír leszabályozott vagy mutált lignin bioszintetikus génjeivel kapcsolatos különböző vizsgálatokról, az ebből eredő hatásokkal a fafeldolgozás hatékonyságára, ha meghatározzák. A lignin bioszintetikus út bármely lépésének aktivitásának csökkentése a PAL-tól a CAD-ig a lignintartalom csökkenését eredményezheti (1.táblázat). Számos paraméter befolyásolja a lignin redukció mértékét, például a célgént és az enzimaktivitás csökkentésének mértékét, ami viszont függ az alkalmazott hangtompító konstrukció hatékonyságától, a géncsalád méretétől és a géncsaládon belüli redundanciától. Általában a C4H-tól a CCR-ig terjedő lépések csökkentése a lignin mennyiségének drámaibb csökkenését eredményezi (Hu et al., 1999; Meyermans et al., 2000; Zhong et al., 2000; Li L. et al., 2003; Jia et al., 2004; Lu et al., 2004; LEPL ++ et al., 2007; Coleman et al., 2008a, b; Bjurhager et al., 2010; Mansfield et al., 2012; Ralph et al., 2012; Min et al., 2014; Van Acker et al., 2014; Zhou et al., 2015, 2018; Saleme et al., 2017; Xiang et al., 2017), mint az F5H, a COMT és a CAD csökkentése (van Doorsselaere et al., 1995; Baucher et al., 1996; Lapierre et al., 1999; Jouanin et al., 2000; Van Acker et al., 2014; Wang et al., 2018). A Lignin redukció az S/G növekedésével járhat, például a C-ben3 ‘ H- (Coleman et al., 2008a; Ralph et al., 2012) és a ccoaomt-downregulated trees (Meyermans et al., 2000), vagy az S/G arány csökkenése, például a CSE – ben (Saleme et al., 2017), és COMT-downregulated fák (van Doorsselaere et al., 1995; Lapierre et al., 1999; Jouanin et al., 2000). Érdekes, hogy az alacsony lignin-4cl-ben szabályozatlan nyárfák S/G növekedése (Min et al., 2014; Xiang et al., 2017), az S/G csökkenése (Voelker et al., 2010; Zhou et al., 2015), vagy a vad típushoz hasonló arányok (Hu et al., 1999; Li L. et al., 2003). Ez a variancia nem társítható a promóterrel vagy a csökkentéshez használt módszerrel, ami arra utal, hogy a némítás mértékének különbségei, növekedési feltételek vagy fejlődési állapot befolyásolja ezt a tulajdonságot. Másrészt a H/G/S lignin összetételre gyakorolt legerősebb hatásokat a C3 ‘ h-ban és a HCT-ben csökkent fák esetében figyelték meg, amelyek a lignint a H egységtartalom jelentős növekedésével rakják le (Coleman et al., 2008a; Ralph et al., 2012; Vanholme et al., 2013a), míg az F5H-t túlexpresszáló fák s egységekben erősen dúsított lignint termelnek (Franke et al., 2000; Li L. et al., 2003; Stewart et al., 2009), és a COMT-ban csökkentett fák drasztikusan csökkentették az egység bioszintézisét (van Doorsselaere et al., 1995; Lapierre et al., 1999).

1. táblázat. A lignin bioszintézis gének módosított expressziójával rendelkező erdei fák áttekintése.
a biomassza feldolgozási hatékonysága a vadon élő növények ligninjében általában kisebb komponenseket képviselő molekulák fokozott beépítésével is módosítható. A ferulinsav beépítése a CCR-hiányos fákba acetálkötések képződését eredményezi a lignin polimerben, amelyek savas biomassza előkezelésekben könnyen hasíthatók (LEPL., 2007; Ralph et al., 2008; Van Acker et al., 2014). Valójában a ferulinsav szintje a ligninben pozitívan korrelált a magasabb szacharifikációs hatékonysággal (Van Acker et al., 2014). Az 5-hidroxikoniferil-alkohol és az 5-hidroxikoniferaldehid beépítése a COMT-hiányos nyárfák ligninjébe (van Doorsselaere et al., 1995; Lapierre et al., 1999; Jouanin et al., 2000; Morreel et al., 2004; Lu et al., 2010) benzodioxán kötéseket hoz létre, amelyek potenciálisan megakadályozzák a lignin és a poliszacharid-hidroxilcsoportok közötti kovalens kapcsolatokat (Weng et al., 2010; Vanholme et al., 2012; Nishimura et al., 2018). Másrészt a COMT-hiány egy kondenzáltabb lignint is eredményez, mivel az S egységfrekvencia csökkenésével viszonylag magasabb a kondenzált blokk-és-5-kötések szintje, valamint az alacsonyabb szintűeké-o-4 kötések. A COMT szempontjából erősen szabályozott nyárfákból származó fa kémiai pépesítése magasabb cellulózhozamot eredményezett, amelyet ellensúlyozott a cellulóz maradék lignintartalma. Ezeknek a fáknak alacsonyabb lignin-és magasabb cellulóz-tartalma volt (Jouanin et al., 2000). Másrészt a COMT-ra szerényen visszafogott nyárfák jelentősen csökkentették a cellulózhozamot, feltehetően azért, mert a lignintartalom normális maradt, míg a ligninnél nagyobb volt a kondenzált kötések gyakorisága, amelyek negatívan befolyásolták a lignin extrakcióját (Lapierre et al., 1999; Pilátus et al., 2002). A cinnamaldehidek beépítése a lignin polimerbe CAD-hiányos fákban rövidebb lignin polimer láncokat eredményez, ezért nagyobb a szabad fenolos végcsoportok aránya, amelyek növelik a polimer oldhatóságát lúg. A cinnamaldehidek beépítése a lignin polimerbe feltehetően csökkenti az alifás lánc kovalens kölcsönhatását a hemicellulózzal, ismét oldhatóbbá téve a lignint. Ezen túlmenően, a kiterjesztett konjugált rendszer miatt, amely akkor keletkezik, amikor egy cinnamaldehidet egy másik monomerrel párosítanak, a beépített cinnamaldehid aromás éterkötése érzékenyebbé válik a lúgos hasításra (Lapierre et al., 1989; Van Acker et al., 2017).
a Lignin polimerizáció egy kombinatorikus radikális kapcsolási folyamat, amely lehetővé teszi a fenolos vegyületek széles skálájának természetes beépítését a lignin polimerbe (Boerjan et al., 2003; Vanholme et al., 2019). A kutatók megkísérelték a lignin mennyiségét és összetételét a biomassza feldolgozásának javítására heterológ gének expressziójával, különböző kompatibilis fenolos vegyületek bioszintézisére és alternatív monolignolként történő beépítésére a lignin polimerbe (Ralph, 2006; Vanholme et al., 2012; Mottiar et al., 2016; Mahon és Mansfield, 2018). Az egyik példa az enzimeket kódoló gének bevezetése, amelyek szükségesek az S egység bioszintéziséhez a pine-ben; az f5h, a COMT és a CAD egyidejű expressziója sikeresen bevezette az S egységeket a Pinus radiata-ban (Wagner et al., 2015; Edmunds et al., 2017). A monolignol 4-o-metiltranszferázt (MOMT4) kódoló gén nyárba történő bevezetése 4-O-metilezett tűlevelű és szinapil-alkoholok képződéséhez vezet, amelyek nem építhetők be a növekvő lignin polimerbe, mert hiányzik belőlük az aromás hidroxilcsoport. Ez a lignin polimerizációjának megállításához vezet, és alacsonyabb lignintartalmú és nagyobb szacharifikációs hatékonyságú fákat eredményez (Bhuiya and Liu, 2010; Cai et al., 2016). A nyárfákat úgy is tervezték, hogy észterkötéseket tartalmazzanak a lignin polimer gerincében. A tűlevelű ferulátum-észtereket egy Feruloil-CoA:Monolignol transzferáz (FMT) gén expressziójával vezettük be a polimerbe Angelica sinensis (Wilkerson et al., 2014), ami a szacharifikáció hatékonyságának javításához vezet különböző előkezelési körülmények között (Wilkerson et al., 2014; Kim et al., 2017; Bhalla et al., 2018), és javult a kraft pulping hatékonysága a vad típushoz képest (Zhou et al., 2017). A monolignol p-kumarát-észtereket nyárfában is kifejlesztették egy rizs p-KUMAROIL-CoA:MONOLIGNOL transzferáz (PMT) gén expressziójával, ami a rezisztens interunit kötések nagyobb gyakoriságát és a G és S terminális egységek nagyobb gyakoriságát eredményezi szabad fenolcsoportokkal (Smith et al., 2015; Sibout et al., 2016). Míg az Arabidopsisban a PMT heterológ expressziója csökkent ligninmennyiséget eredményezett, amelyet fokozott szacharifikációs hatékonyság kísér (Sibout et al., 2016), a nyárfában nem csökkent a lignin mennyisége, és a szacharifikációs hatékonyságot nem határozták meg (Smith et al., 2015).
míg a lignin mennyiségének és összetételének számos módosítása javulást mutatott a biomassza feldolgozásában, ezeket a módosításokat gyakran a biomassza hozamának büntetése kísérte (LEPL)., 2007; Wadenboxcock et al., 2008; Wagner et al., 2009; Voelker et al., 2010; Stout et al., 2014; Van Acker et al., 2014; Sykes et al., 2015; Zhou et al., 2018). Egy nemrégiben végzett metasztázis zavart 21 lignin bioszintézis gének P-ben. trichocarpa, és átfogóan integrálta a 221 vonal transzkriptomikus, proteomikus, fluxomikus és fenomikus adatainak eredményeit. A szerzők arra a következtetésre jutottak, hogy a fa növekedése nem kapcsolódik a lignin mennyiségéhez, az alegység összetételéhez vagy a specifikus kapcsolatokhoz (Wang et al., 2018), hanem inkább korrelált az összeomlott xylem hajók jelenlétével (Coleman et al., 2008a, b; Wagner et al., 2009; Voelker et al., 2010; Vargas et al., 2016; de Meester et al., 2018), a sejtfal integritási útjának aktiválása (Bonawitz et al., 2014) és / vagy a kémiai inhibitorok felhalmozódása (Gallego-Giraldo et al., 2011; Muro-Villanueva et al., 2019).
míg jelentős erőfeszítéseket tettek a lignin tartalom csökkentésére a lignin bioszintetikus gének csökkentése révén, a lignin útvonalának felszabályozásáról és a lignin túltermeléséről kevés tanulmány készült. Valójában az F5H túlzott expressziójáról szóló jelentések a lignintartalom változatlan vagy akár csökkenését mutatják (Huntley et al., 2003; Li L. et al., 2003; Stewart et al., 2009; Mansfield et al., 2012; Edmunds et al., 2017). A CAD és a COMT túlexpressziója inkább géncsendesítést eredményezett, mint upregulációt, vagy az expressziós szintekre gyakorolt hatást nem észleltek (Baucher et al., 1996; Lapierre et al., 1999; Jouanin et al., 2000; LEPL ++ et al., 2007; Van Acker et al., 2014). Az R2r3-MYB transzkripciós faktorok PtoMYB92, PtoMYB216 és PtoMYB74 túlexpressziója további xilem rétegeket, vastagabb xilem sejtfalakat, valamint méhen kívüli lignin lerakódást eredményezett ,és a növények 13-50% – kal több lignint halmoztak fel (Tian Q. et al., 2013; Li C. F. et al., 2015; Li et al., 2018). A MYB túlexpressziós vonalak konstitutív módon szabályozták a lignin bioszintézis útvonal génjeit, és míg a myb92-t és a MYB74-et túlexpresszáló növények biomassza-büntetést kaptak, a MYB216 túlexpressziója akár 50% – kal több lignint és fejlődési fenotípust nem eredményezett. Mivel a lignint egyre inkább a vegyi anyagok fenntartható termelésének fontos erőforrásának tekintik (Cao et al., 2018) tovább kell vizsgálni a lignint túltermelő növények tervezését.
terepi vizsgálatok
a fent tárgyalt példák egyértelműen azt mutatják, hogy a lignin-tervezés a fenilpropanoid útvonal gének-vagy heterológ gének expressziója – révén növelheti a lignocellulóz biomassza feldolgozási hatékonyságát. Gyakorlati és szabályozási okokból a legtöbb tanulmány az üvegházban termesztett fák elemzéséből származó adatokról számol be. Az üvegházban termesztett fákkal végzett kísérletek azonban általában nem veszik figyelembe a fejlődési folyamatokat, mint például a növekedés abbahagyása és a nyugalmi állapot. Ezenkívül az üvegházhatású kísérletek nem nyújtanak elegendő betekintést a tervezett növény kölcsönhatásába olyan környezeti tényezőkkel, mint a talajtípus, a szél és a kórokozók. Ezen interakciók megértése fontos lépés a kutatási eredmények kereskedelmi alkalmazások felé történő fordításában. Valójában az olyan tanulmányok által készített munka, amelyekre engedélyt adtak a terepi kísérletek létrehozására, kiemeli az üvegházhatású és a szántóföldi fák fenotípusának fontos különbségeit. Az 1.táblázat összefoglalja a 4cl, CCoAOMT, CCR, COMT és CAD csökkentett szabályozású fákkal végzett terepi vizsgálatokról szóló jelentéseket.
megerősítve a módosított lignocellulóz biomassza szubsztrátként való alkalmazásának lehetőségét, számos ligninnel tervezett terepi kísérletben termesztett fa javulást mutatott a fafeldolgozásban. Nyárfa downregulated a CCoAOMT nőtt 5 éve egy terepi vizsgálat Pekingben (Kína), azt mutatta, fokozott glükóz és xilóz kiadás után cukrosítás (Wang et al., 2012). A CCR-re csökkentett nyárfa, amelyet egy franciaországi terepi kísérlet során termesztettek, alkalmasabbnak bizonyult a kémiai kraft-pépesítésre (LEPL ++ et al., 2007). Két további, Franciaországban és Belgiumban, CCR-rel lefelé szabályozott nyárfával végzett terepi vizsgálat az etanol előállításának akár 160% – os javulását eredményezte egyidejű szacharifikációs és fermentációs (SSF) vizsgálatokban; a növények azonban akár 50% – kal csökkentették a biomasszát (Van Acker et al., 2014). A CAD-downregulated nyárral végzett terepi kísérletek szintén ígéretes eredményeket mutattak. Ezek a fák valamivel kevesebb lignint mutattak, mint a vadon élő fajok, és alkalmasabbnak bizonyultak a kraft delignification (Lapierre et al., 1999). A Franciaországban és az Egyesült Királyságban végzett nagyobb léptékű terepkísérletek során termesztett vonalak következetesen a lignin mennyiségének enyhe csökkenését és a kraft pépesítésének kereskedelmileg relevánsnak ítélt javulását mutatták, mivel a növényeknek 6%-kal kevesebb lúgra volt szükségük ahhoz, hogy a vadon élő fákhoz hasonló deligifikációt érjenek el (Pilate et al., 2002).
azonban mind a biomassza hozamáról, mind a későbbi feldolgozás hatékonyságáról szóló ellentmondásos jelentések azt sugallják, hogy ezeket a paramétereket nagymértékben befolyásolják a környezeti tényezők. Míg Kínában a 4cl-es lefelé szabályozott nyárfákkal végzett terepi vizsgálat megállapította, hogy a lignintartalom 28% – os csökkenése mellett a vad típushoz képest a fák magassága körülbelül 8% – kal nőtt (Tian X. M. et al., 2013), összhangban az üvegházhatású tanulmányokkal (Hu et al., 1999), más terepi kísérletek azt találták, hogy a 4CL-ben lefelé szabályozott nyárfák csökkentették a biomasszát, sőt néha eltörpültek (Voelker et al., 2010; Stout et al., 2014; Marchin et al., 2017). A jelentések eltérnek az ezekből a 4cl-ből származó, lefelé szabályozott szántóföldi nyárfákból származó fa downstream feldolgozási hatékonyságáról is. Míg a cukor visszanyerésének akár 100% – os növekedését találták az 4CL1-downregulated fáknál (35S-vezérelt antiszensz 4CL konstrukció), amelyet az Egyesült Államok hegyi helyén termesztettek (Xiang et al., 2017), az Oregonban (Egyesült Államok) végzett terepi vizsgálatokból nyert adatok azt találták, hogy a Pt4CL1 promoter által vezérelt antiszensz elhallgattatott 4CL1 nyárfák nem javultak a szacharifikációs hatékonyságban a vad típushoz képest (Voelker et al., 2010). Hasonlóképpen, egy hosszú távú tanulmány Wenlingben (Kína) megállapította, hogy a 4CL-vel lefelé szabályozott nyárfák nem mutattak jelentős javulást a cukorhozamban a vad típushoz képest (Wang et al., 2012). Mindkét utóbbi esetben a fák enyhe csökkenést mutattak a lignin mennyiségében, ami nem eredményezett nagyobb feldolgozási hatékonyságot, potenciálisan az extraktumok magasabb koncentrációja miatt, amelyek zavarhatják az enzimatikus aktivitást (Voelker et al., 2010).
terepi vizsgálatok kimutatták, hogy a környezeti tényezők befolyásolhatják a ligifikációt és visszaállíthatják a vad típusú tulajdonságokat azokhoz a szintekhez képest, amelyeket ugyanazon növények üvegházban történő termesztésekor értek el. Míg a 4cl-vel csökkentett fák lignintartalma csökkent, ha üvegházi körülmények között termesztették, ugyanazon 4cl-es antiszensz nyárfák elemzése, de a szántóföldön termesztették, gyakran kimutatta, hogy a lignintartalom megnőtt az üvegházban termesztett fákhoz képest, sőt néha vad típusú szintre is visszaállt (Stout et al., 2014; Xiang et al., 2017). Hasonlóképpen, a lignin szintje sokkal kevésbé csökkent a CCR-hiányos nyárfákban, amikor a szántóföldön termesztették őket, mint amikor az üvegházban termesztették őket (Van Acker et al., 2014). Legalább a CCR-hiányos nyárfák esetében lehetséges, hogy a szántóföldön termesztett fák Magasabb ligifikációs szintje annak köszönhető, hogy a fa mintákat télen vették. Amikor a fa növekedése ősszel megszűnik, a fáknak még van idejük sejtfaluk teljes ligifikálására, mire a fa nyugalmi állapotba kerül, összehasonlítva az üvegházban termesztett fákkal, amelyek folyamatosan új xilemet fejlesztenek ki. Kimutatták, hogy a Lignin összetétele is különbözik az üvegházban termesztett és a szántóföldön termesztett alacsony lignin fák között. 4cl-az észak-karolinai mezőn termesztett, lefelé szabályozott nyárfák ligninje alacsonyabb S/G volt, mint amikor ugyanazokat a vonalakat termesztették az üvegházban (Stout et al., 2014).
ezek az eredmények együttesen azt mutatják, hogy az üvegházban termesztett fákból nyert adatokat nem lehet könnyen extrapolálni a szántóföldön termesztett fákra, ami alátámasztja a különböző helyszíneken végzett terepi kísérletek szükségességét. Egyes sorok hozambüntetést mutattak be, ami kevésbé érdekessé tette őket az alkalmazások számára, kiemelve a hozambüntetés molekuláris alapjának jobb megértésének szükségességét, valamint a probléma leküzdésére irányuló stratégiák kidolgozását.
kimutatták, hogy a Lignin fontos szerepet játszik a kórokozókkal szembeni rezisztenciában (Miedes et al., 2014; Zhao és Dixon, 2014), és kulcsfontosságú szerepet játszik abban, hogy az üzem vizet szállítson. Ez arra utal, hogy a lignin-módosítások hatással lehetnek a növényi stressztűrésre. Bár további vizsgálatokra van szükség ennek a lehetőségnek a teljes körű kezeléséhez, a 4cl, a COMT és a CAD nyárban történő csökkentése nem változtatta meg drámai módon a levelekkel táplálkozó növényevők táplálkozási teljesítményét (Tiimonen et al., 2005; Brodeur-Campbell et al., 2006; Hjalten et al., 2013). A COMT és a CAD nyárban történő csökkentésének hatását a növény-rovar kölcsönhatásokra a szántóföldön termesztett fákon is értékelték, és kimutatták, hogy a ligninnel módosított fák normális előfordulási gyakorisággal látogatják és táplálják a rovarokat, valamint normális válaszokat adnak a mikrobiális kórokozókra (Pilate et al., 2002; Halpin et al., 2007). Ezek az eredmények azt mutatják, hogy a módosított lignint tartalmazó fák nem feltétlenül szenvednek többet a kártevőktől és betegségektől, mint a vadon élő növények. Ennek ellenére az endoszféra bakteriális mikrobiomjának profilozása a szántóföldön termesztett, CCR-ben lefelé szabályozott nyárfákból betakarított fa eltolódását mutatta a baktériumközösségben, feltehetően a xilemben lévő fenolos metabolitok megváltozott bősége miatt (Beckers et al., 2017).
figyelembe véve a lignin szerepét a xilem működésében és szerkezetében, néhány alacsony ligninnel módosított nyárfa vízviszonyait értékelték. 4cl-downregulated nyár kiderült, hogy csökkent a hidraulikus vezetőképesség, potenciálisan zavarja a növény növekedését (Marchin et al., 2017). Hidraulikus stressz kísérletek nyárfa downregulated CCR, COMT vagy CAD azt mutatta, hogy ezek a növények volt egy kisebb ellenállás kavitáció, miközben a normál Xylem hidraulikus vezetőképesség és a vízi közlekedés (Awad et al., 2012). Ezek az eredmények arra utalnak, hogy az alacsony lignin mutánsok növekedését befolyásolhatja a víz rendelkezésre állása. Mint minden klasszikus tenyésztésből származó új hibrid esetében, terepi tesztekre van szükség a ligninnel módosított fák terepi teljesítményének és stressztűrésének értékeléséhez.
A Lignintechnika kilátásai az erdei fákban
a lignintermelő növények teljesítményét úgy tűnik, hogy nagymértékben befolyásolják a környezeti feltételek. Nem világos azonban, hogy az üvegházban termesztett és a szántóföldön termesztett fák, vagy a különböző mezőn termesztett fák között megfigyelt különbségek a génszuppresszió különböző szintjeiből vagy a módosított tulajdonságnak a környezettel való kölcsönhatásából származnak-e (GxE). Valójában az instabil downreguláció az rnai-n alapuló géncsendesítési technikák hiányossága. Ennek tanúja a vörös xilem fenotípus változásának megfigyelése, amely akkor figyelhető meg, amikor bizonyos lignin bioszintézis gének, például CAD, COMT, vagy CCR, le vannak szabályozva. A vörös xilém színe gyakran nem egyenletes az egész xilémben, hanem olyan foltokban jelenik meg, amelyek a géncsendesítés változó szintjét tükrözik (LEPL)., 2007; Voelker et al., 2010; Van Acker et al., 2014; 2. ábra). Ezenkívül a géncsendesítési módszerek alkalmazása potenciálisan a közeli rokon géncsalád tagjainak egyidejű elnémítását eredményezheti – talán különböző mértékben -, elhomályosítva az értelmezéseket és álcázva az egyes gének downregulációjának hatásait.

2. ábra. Foltos gén downregulation által RNAi. Foltos vörös xilem fenotípus a CCR-hiányos nyárfák törzsén (jobbra), amelyet egy belgiumi terepi kísérlet során termesztettek. A piros xilem a CCR downreguláció területeit jelzi. A vad típusú fákból származó fa fehéres (balra).
ezek a kérdések most már könnyen leküzdhetők CRISPR-alapú génszerkesztési technológiák alkalmazásával, amelyek lehetővé teszik a stabil funkcióvesztés mutációk (knock-out) specifikus célgénekben, lehetővé téve az egyes gének funkciójának boncolását a családokon belül. Például az egyes 4cl géncsaládok megcélzása a nyárban azt mutatta, hogy a 4cl1 összefügg a ligifikációval, míg a 4CL2 részt vesz a proanthocyanidin termelésében (Zhou et al., 2015). A knock-out allélek mellett a CRISPR-alapú génszerkesztés új allélok létrehozását is lehetővé teszi, amelyek az enzimaktivitás részleges csökkenését eredményezik. Ez lehetővé teszi a maradék enzimaktivitás szintjének finomhangolását és a gyakran megfigyelhető hozambüntetés megkerülését, amikor a lignin mennyisége egy küszöbszint alá csökken. Egy másik ígéretes út lignin engineering erdei fák lehetővé tette a CRISPR-alapú Genom engineering egyidejű szerkesztése több gén (allél egymásra), hogy optimalizálja a biomassza feldolgozás hatékonyságát, mint példázza Arabidopsis ahol egymásra a transzaldoláz (tra) és comt mutációk, a c4h és comt mutációk, vagy a 4cl és comt mutációk eredményezett additív és szinergikus javulást szacharifikációs hatékonyság (de Vries et al., 2018). Valóban, egy rendszer megközelítés P. trichocarpa azt jósolja, hogy a PAL és a CCoAOMT, vagy a PAL, a C3 ‘ H és a CCOAOMT egyidejű downregulációja jelentősen javítja a fa tulajdonságait és a cukor felszabadulását (Wang et al., 2018).
a CRISPR-alapú genomszerkesztés használata a cellulóz-és papíripar, valamint a biofinomító iparágak fajavításában, valamint a lignin hidrogénolitikus lebontásából származó platformaromatikumok előállításához akkor lesz a legértékesebb, ha ezt a technológiát stratégiailag kombinálják más tenyésztési technikákkal (3.ábra). Valójában a lignin mennyiségében és S/G összetételében már nagy eltérések vannak az erdei fák természetes populációiban (Studer et al., 2011). Tekintettel arra, hogy mindkét tulajdonság befolyásolja a glükóz felszabadulását szacharifikációkor (Yoo et al., 2018), kihasználva ezt a genetikai sokféleséget hagyományos tenyésztéssel, genome wide Association Studies (GWAS) segítségével (Porth et al., 2013; Fahrenkrog et al., 2017; Liu et al., 2018), tenyésztés ritka hibás allélokkal (BRDA) (Vanholme et al., 2013a) vagy genomikus szelekció (Yin et al., 2010; Muchero et al., 2015; Pawar et al., 2018; Xie et al., 2018), értékes stratégia olyan vonalak beszerzésére, amelyek javították a fafeldolgozás hatékonyságát. Miután az elit fákat ezekkel a tenyésztési módszerekkel megszerezték, a géntechnológia és a specifikus gének CRISPR-alapú génszerkesztése nagyon ígéretes út ezen elit genotípusok további fejlesztésére anélkül, hogy genetikai felépítésüket megszakítanák, és hosszú tenyésztési ciklusokon mennének keresztül. Tekintettel a közelgő éghajlati összeomlásra, nincs több időnk elvesztegetni ezeket az új tenyésztési technikákat a biobázisú gazdaság versenyében.
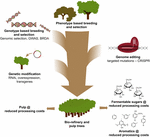
3. ábra. Az erdei fák genetikai javítása tenyésztési eszközök kombinációjával. Az erdei fák genetikai fejlesztésének felgyorsítása a cellulóz és a biofinomító alkalmazásokhoz, a klasszikus és az új tenyésztési eszközöket okosan kell kombinálni. A klasszikus tenyésztés magában foglalja a fák fenotípusos kiválasztását ellenőrzött keresztekhez, majd fenotípusos szelekció. Az advent a genom szekvencia információ sok erdei fák, új stratégiák, mint a genom szelekció, Genome wide Association Studies (GWAS) és a tenyésztés ritka hibás allélokkal (BRDA) fejlesztettek ki, hogy gyorsítsák fel a befogását és dúsítása DNS polimorfizmusok társított előnyös tulajdonságok. A CRISPR-alapú genomszerkesztés lehetővé teszi a genom módosítását oly módon, hogy utánozza a természetes polimorfizmusokat. A genetikai módosítás magában foglalja az idegen DNS stabil integrációját a fába az (An) enzim(ek) túltermelése vagy az (a) gén(ek) alulszabályozása érdekében. A klasszikus és az új tenyésztési technikák ötvözése szükséges ahhoz, hogy a társadalom számára megfelelő minőségű fát biztosítsunk.
szerzői hozzájárulások
minden felsorolt szerző jelentős, közvetlen és szellemi hozzájárulást nyújtott a műhöz, és jóváhagyta a közzétételt.
finanszírozás
elismerjük az IWT-SBO bioleum projekt (130039.számú támogatás), valamint az SBO-FISH részleges finanszírozását az ARBOREF projekten keresztül. Az AC az FWO-tól és az Európai Unió Horizont 2020 kutatási és Innovációs programjától kapott támogatást a Marie SK 665501.sz. Az LdV – t az Institute for the promotion of Innovation through Science and Technology in Flanders (IWT-Vlaanderen) finanszírozta egy predoctoral ösztöndíj céljából.
összeférhetetlenségi nyilatkozat
a szerzők kijelentik, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végezték, amelyek potenciális összeférhetetlenségnek tekinthetők.
köszönetnyilvánítások
köszönjük Annick Bleysnek, hogy elkészítette ezt a kéziratot benyújtásra.
Boerjan, W., Ralph, J., and Baucher, M. (2003). Lignin bioszintézis. Annu. Rev. Növény Biol. 54, 519–546.
PubMed Abstract/Google Scholar
Lapierre, C., Pollet, B., Petit-Conil, M., Toval, G., Romero, J., Pilátus, G., et al. (1999). A ligninek szerkezeti változásai a transzgenikus nyárfákban depressziós cinnamil-alkohol-dehidrogenáz vagy koffeinsav O-metiltranszferáz aktivitással ellentétes hatással vannak az ipari nátronpépezés hatékonyságára. Növényi Fiziol. 119, 153–163.
PubMed absztrakt / Google Tudós
Ralph, J. (2006). “Mi teszi a jó monolignol helyettesítőt?, “In The Science and Lore of the Plant Cell Wall-bioszintézis, szerkezet és funkció, Szerk. T. Hayashi (Boca Raton, FL: BrownWalker Press).
Google Scholar