Obiettivi formativi
Alla fine di questa sezione, potrai:
- Spiegare i processi di glicolisi
- Descrivere il percorso di una molecola di piruvato attraverso il ciclo di Krebs
- Spiega il trasporto di elettroni attraverso la catena di trasporto degli elettroni
- Descrivere il processo di produzione di ATP attraverso la fosforilazione ossidativa
- Riassumere il processo di gluconeogenesi
I carboidrati sono molecole organiche composto di carbonio, idrogeno e atomi di ossigeno. La famiglia di carboidrati comprende sia zuccheri semplici che complessi. Glucosio e fruttosio sono esempi di zuccheri semplici e amido, glicogeno e cellulosa sono tutti esempi di zuccheri complessi. Gli zuccheri complessi sono anche chiamati polisaccaridi e sono costituiti da più molecole di monosaccaridi. I polisaccaridi servono come accumulo di energia (ad esempio, amido e glicogeno) e come componenti strutturali (ad esempio, chitina negli insetti e cellulosa nelle piante).
Durante la digestione, i carboidrati vengono suddivisi in zuccheri semplici e solubili che possono essere trasportati attraverso la parete intestinale nel sistema circolatorio per essere trasportati in tutto il corpo. La digestione dei carboidrati inizia in bocca con l’azione dell’amilasi salivare sugli amidi e termina con i monosaccaridi assorbiti attraverso l’epitelio dell’intestino tenue. Una volta che i monosaccaridi assorbiti vengono trasportati nei tessuti, inizia il processo di respirazione cellulare (Figura 1). Questa sezione si concentrerà prima sulla glicolisi, un processo in cui il glucosio monosaccaride viene ossidato, rilasciando l’energia immagazzinata nei suoi legami per produrre ATP.
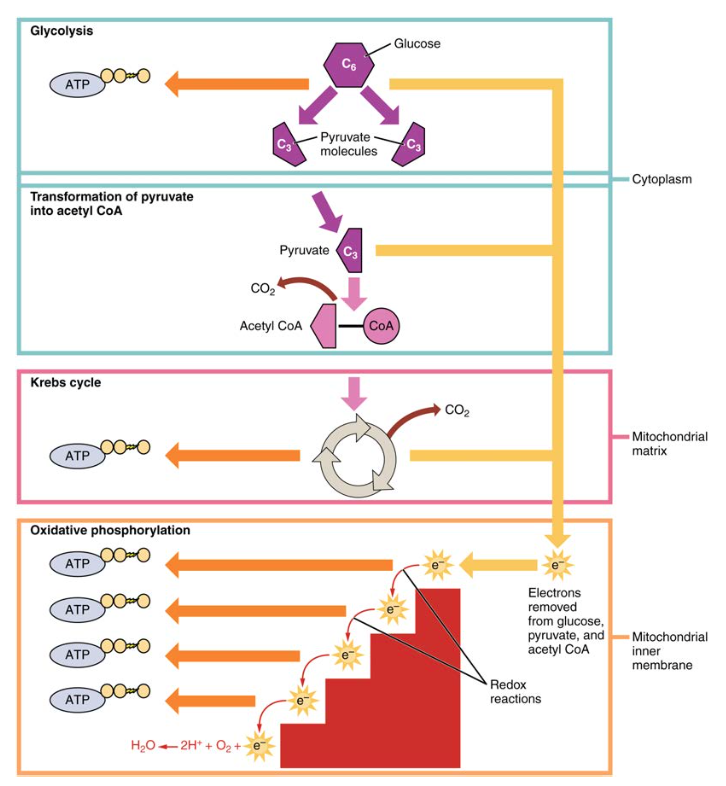
Figura 1. La respirazione cellulare ossida le molecole di glucosio attraverso la glicolisi, il ciclo di Krebs e la fosforilazione ossidativa per produrre ATP.
Glicolisi
Il glucosio è la fonte di energia più facilmente disponibile del corpo. Dopo che i processi digestivi rompono i polisaccaridi in monosaccaridi, incluso il glucosio, i monosaccaridi vengono trasportati attraverso la parete dell’intestino tenue e nel sistema circolatorio, che li trasporta al fegato. Nel fegato, gli epatociti passano il glucosio attraverso il sistema circolatorio o immagazzinano il glucosio in eccesso come glicogeno. Le cellule del corpo assorbono il glucosio circolante in risposta all’insulina e, attraverso una serie di reazioni chiamate glicolisi, trasferiscono parte dell’energia in glucosio all’ADP per formare ATP (Figura 2). L’ultimo passo nella glicolisi produce il prodotto piruvato.
La glicolisi inizia con la fosforilazione del glucosio da parte dell’esochinasi per formare glucosio-6-fosfato. Questo passaggio utilizza un ATP, che è il donatore del gruppo fosfato. Sotto l’azione della fosfofruttochinasi, il glucosio-6-fosfato viene convertito in fruttosio-6-fosfato. A questo punto, un secondo ATP dona il suo gruppo fosfato, formando fruttosio-1,6-bisfosfato. Questo zucchero a sei atomi di carbonio è diviso per formare due molecole fosforilate a tre atomi di carbonio, gliceraldeide-3-fosfato e diidrossiacetone fosfato, che sono entrambi convertiti in gliceraldeide-3-fosfato. La gliceraldeide-3-fosfato è ulteriormente fosforilata con gruppi donati dal diidrogeno fosfato presente nella cellula per formare la molecola a tre atomi di carbonio 1,3-bisfosfoglicerato. L’energia di questa reazione deriva dall’ossidazione di (rimozione di elettroni da) gliceraldeide-3-fosfato. In una serie di reazioni che portano al piruvato, i due gruppi fosfatici vengono quindi trasferiti a due ADP per formare due ATP. Pertanto, la glicolisi utilizza due ATPS ma genera quattro ATPS, producendo un guadagno netto di due ATPS e due molecole di piruvato. In presenza di ossigeno, il piruvato continua al ciclo di Krebs (chiamato anche ciclo dell’acido citrico o ciclo dell’acido tricarbossilico (TCA), dove viene estratta e trasmessa energia aggiuntiva.
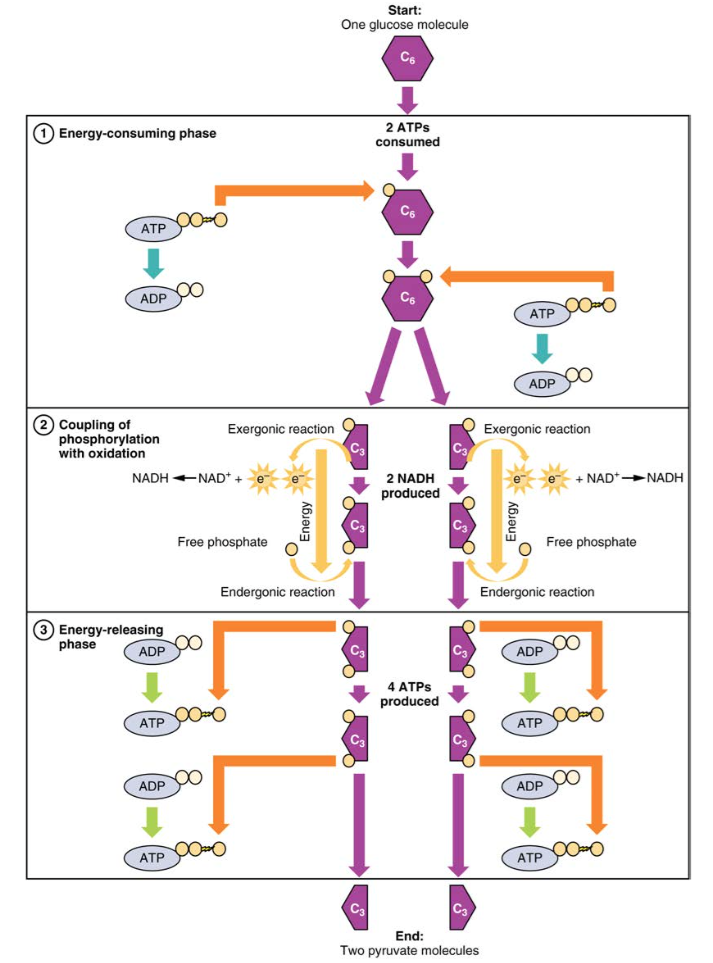
Figura 2. Durante la fase di consumo energetico della glicolisi, vengono consumati due ATP, trasferendo due fosfati alla molecola di glucosio. La molecola di glucosio si divide quindi in due composti a tre atomi di carbonio, ciascuno contenente un fosfato. Durante la seconda fase, un fosfato aggiuntivo viene aggiunto a ciascuno dei composti a tre atomi di carbonio. L’energia per questa reazione endergonica è fornita dalla rimozione (ossidazione) di due elettroni da ciascun composto a tre atomi di carbonio. Durante la fase di rilascio di energia, i fosfati vengono rimossi da entrambi i composti a tre atomi di carbonio e utilizzati per produrre quattro molecole di ATP.
Guarda questo video per conoscere la glicolisi:
La glicolisi può essere divisa in due fasi: consumo di energia (chiamato anche innesco chimico) e rendimento energetico. La prima fase è la fase che consuma energia, quindi richiede due molecole di ATP per iniziare la reazione per ogni molecola di glucosio. Tuttavia, la fine della reazione produce quattro ATPS, con conseguente guadagno netto di due molecole di energia ATP.
La glicolisi può essere espressa come la seguente equazione:
\text{Glucose}+2\text{ATP}+2\text{NAD}^{+}+4\text{ADP}+2\text{P}_{i}\to\text{Pyruvate}+4\text{ATP}+2\text{NADH}+2\text{H}^{+}
Questa equazione afferma che il glucosio, in combinazione con ATP (la fonte di energia), NAD+ (un coenzima che serve come accettore di elettroni) e fosfato inorganico, si scompone in due molecole di piruvato, generando quattro molecole di ATP per un rendimento netto del due di ATP e due energetiche contenenti coenzimi NADH. Il NADH che viene prodotto in questo processo verrà utilizzato in seguito per produrre ATP nei mitocondri. È importante sottolineare che, alla fine di questo processo, una molecola di glucosio genera due molecole di piruvato, due molecole di ATP ad alta energia e due molecole di NADH che trasportano elettroni.
Le seguenti discussioni sulla glicolisi includono gli enzimi responsabili delle reazioni. Quando il glucosio entra in una cellula, l’enzima esochinasi (o glucochinasi, nel fegato) aggiunge rapidamente un fosfato per convertirlo in glucosio-6-fosfato. Una chinasi è un tipo di enzima che aggiunge una molecola di fosfato a un substrato (in questo caso, glucosio, ma può essere vero anche per altre molecole). Questa fase di conversione richiede un ATP ed essenzialmente intrappola il glucosio nella cellula, impedendogli di passare attraverso la membrana plasmatica, consentendo così la glicolisi di procedere. Funziona anche per mantenere un gradiente di concentrazione con livelli di glucosio più elevati nel sangue rispetto ai tessuti. Stabilendo questo gradiente di concentrazione, il glucosio nel sangue sarà in grado di fluire da un’area ad alta concentrazione (il sangue) in un’area a bassa concentrazione (i tessuti) da utilizzare o immagazzinare. L’esochinasi si trova in quasi tutti i tessuti del corpo. La glucochinasi, d’altra parte, è espressa in tessuti che sono attivi quando i livelli di glucosio nel sangue sono alti, come il fegato. Esochinasi ha una maggiore affinità per il glucosio rispetto glucochinasi e quindi è in grado di convertire il glucosio ad un tasso più veloce di glucochinasi. Questo è importante quando i livelli di glucosio sono molto bassi nel corpo, in quanto consente al glucosio di viaggiare preferenzialmente a quei tessuti che lo richiedono di più.
Nella fase successiva della prima fase della glicolisi, l’enzima glucosio-6-fosfato isomerasi converte il glucosio-6-fosfato in fruttosio-6-fosfato. Come il glucosio, il fruttosio è anche uno zucchero contenente sei carbonio. L’enzima fosfofruttochinasi – 1 aggiunge quindi un altro fosfato per convertire il fruttosio-6-fosfato in fruttosio-1-6-bisfosfato, un altro zucchero a sei atomi di carbonio, usando un’altra molecola di ATP. Aldolasi poi rompe questo fruttosio-1-6-bifosfato in due molecole a tre atomi di carbonio, gliceraldeide-3-fosfato e diidrossiacetone fosfato. L’enzima triosefosfato isomerasi converte quindi il fosfato diidrossiacetone in una seconda molecola di gliceraldeide-3-fosfato. Pertanto, entro la fine di questa fase di innesco chimico o di consumo energetico, una molecola di glucosio viene scomposta in due molecole di gliceraldeide-3-fosfato.
La seconda fase della glicolisi, la fase che produce energia, crea l’energia che è il prodotto della glicolisi. La gliceraldeide-3-fosfato deidrogenasi converte ogni gliceraldeide-3-fosfato a tre atomi di carbonio prodotto durante la fase di consumo energetico
in 1,3-bifosfoglicerato. Questa reazione rilascia un elettrone che viene poi raccolto da NAD+ per creare una molecola di NADH. NADH è una molecola ad alta energia, come l’ATP, ma a differenza dell’ATP, non viene utilizzata come valuta energetica dalla cellula. Poiché ci sono due molecole di gliceraldeide-3-fosfato, due molecole di NADH vengono sintetizzate durante questa fase. Ogni 1,3-bisfosfoglicerato viene successivamente defosforilato (cioè un fosfato viene rimosso) dalla fosfoglicerato chinasi in 3-fosfoglicerato. Ogni fosfato rilasciato in questa reazione può convertire una molecola di ADP in una molecola di ATP ad alta energia, con un conseguente guadagno di due molecole di ATP.
L’enzima fosfoglicerato mutasi converte quindi le molecole di 3-fosfoglicerato in 2-fosfoglicerato. L’enzima enolasi agisce quindi sulle molecole di 2-fosfoglicerato per convertirle in molecole di fosfoenolpiruvato. L’ultima fase della glicolisi comporta la defosforilazione delle due molecole di fosfoenolpiruvato da parte della piruvato chinasi per creare due molecole di piruvato e due molecole di ATP.
In sintesi, una molecola di glucosio si scompone in due molecole di piruvato e crea due molecole di ATP netto e due molecole di NADH mediante glicolisi. Pertanto, la glicolisi genera energia per la cellula e crea molecole di piruvato, che possono essere ulteriormente elaborati attraverso l’aerobica ciclo di Krebs (chiamato anche ciclo dell’acido citrico o ciclo dell’acido tricarbossilico); convertito in acido lattico o alcol (nel lievito di birra) da fermentazione; o successivamente utilizzati per la sintesi di glucosio attraverso la gluconeogenesi.
Respirazione anaerobica
Quando l’ossigeno è limitato o assente, il piruvato entra in una via anaerobica. In queste reazioni, il piruvato può essere convertito in acido lattico. Oltre a generare un ATP aggiuntivo, questa via serve a mantenere bassa la concentrazione di piruvato, quindi la glicolisi continua e ossida il NADH nel NAD + necessario per la glicolisi. In questa reazione, l’acido lattico sostituisce l’ossigeno come accettore finale di elettroni. La respirazione anaerobica si verifica nella maggior parte delle cellule del corpo quando l’ossigeno è limitato o i mitocondri sono assenti o non funzionali. Ad esempio, poiché gli eritrociti (globuli rossi) mancano di mitocondri, devono produrre il loro ATP dalla respirazione anaerobica. Questo è un percorso efficace di produzione di ATP per brevi periodi di tempo, che vanno da secondi a pochi minuti. L’acido lattico prodotto si diffonde nel plasma e viene trasportato nel fegato, dove viene riconvertito in piruvato o glucosio attraverso il ciclo Cori. Allo stesso modo, quando una persona esercita, i muscoli usano ATP più velocemente dell’ossigeno che può essere consegnato a loro. Dipendono dalla glicolisi e dalla produzione di acido lattico per una rapida produzione di ATP.
Respirazione aerobica
In presenza di ossigeno, il piruvato può entrare nel ciclo di Krebs dove viene estratta energia aggiuntiva mentre gli elettroni vengono trasferiti dal piruvato ai recettori NAD+, GDP e FAD, con l’anidride carbonica come “prodotto di scarto” (Figura 3). Il NADH e il FADH2 passano elettroni alla catena di trasporto degli elettroni, che utilizza l’energia trasferita per produrre ATP. Come il punto terminale nella catena di trasporto dell’elettrone, l’ossigeno è l’accettore terminale dell’elettrone e crea l’acqua dentro i mitocondri.
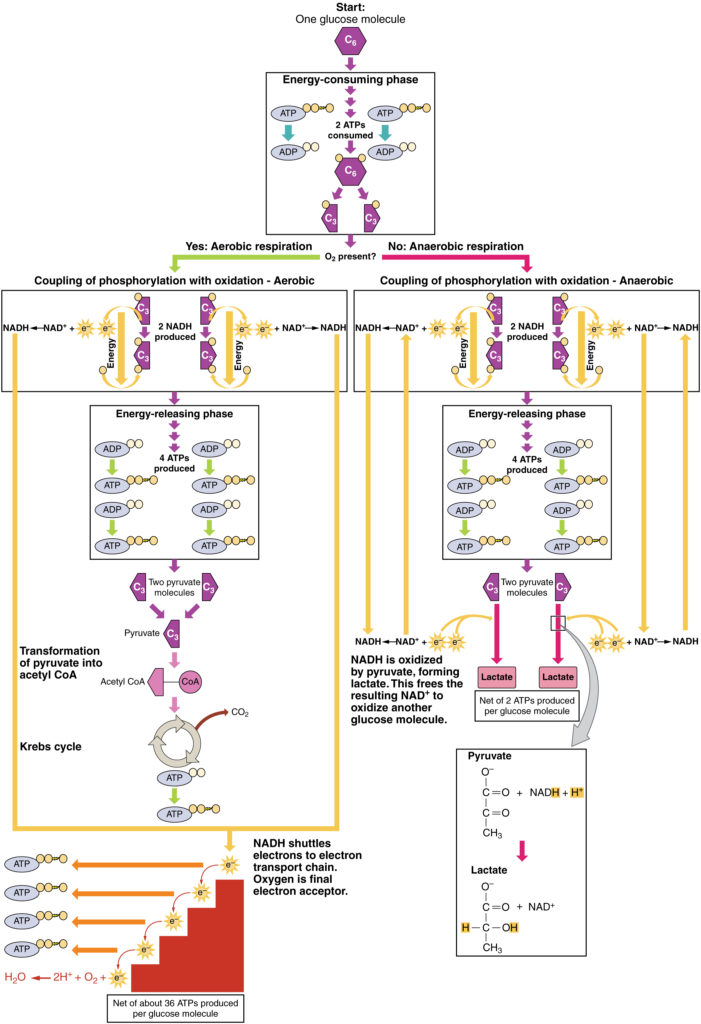
Figura 3. Fare clic per visualizzare un’immagine ingrandita. Il processo di respirazione anaerobica converte il glucosio in due molecole di lattato in assenza di ossigeno o all’interno di eritrociti privi di mitocondri. Durante la respirazione aerobica, il glucosio viene ossidato in due molecole di piruvato.
Ciclo di Krebs/Ciclo dell’acido citrico / Ciclo dell’acido tricarbossilico
Le molecole di piruvato generate durante la glicolisi vengono trasportate attraverso la membrana mitocondriale nella matrice mitocondriale interna, dove vengono metabolizzate dagli enzimi in una via chiamata ciclo di Krebs (Figura 4). Il ciclo di Krebs è anche comunemente chiamato ciclo dell’acido citrico o ciclo dell’acido tricarbossilico (TCA). Durante il ciclo di Krebs, vengono create molecole ad alta energia, tra cui ATP, NADH e FADH2. NADH e FADH2 passano quindi elettroni attraverso la catena di trasporto degli elettroni nei mitocondri per generare più molecole di ATP.
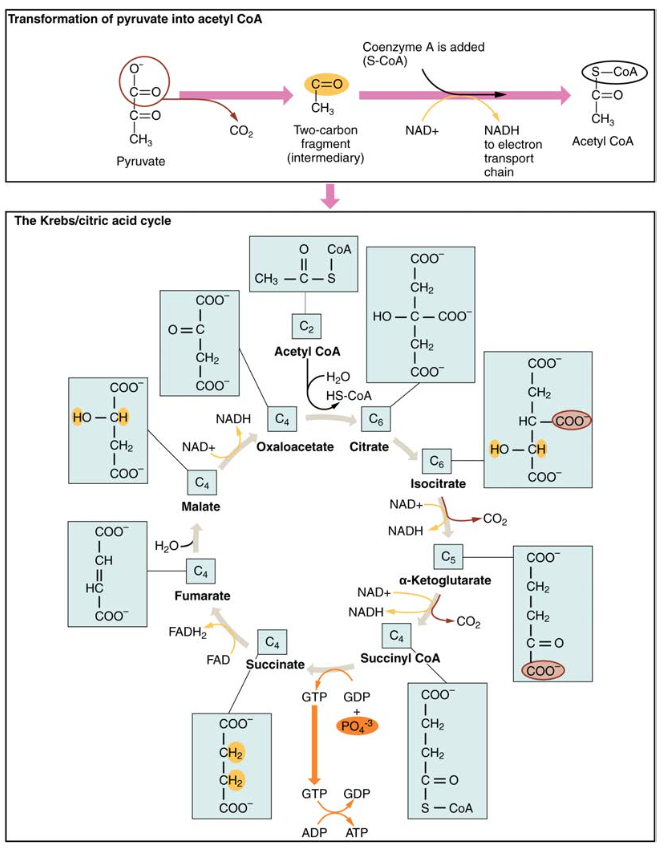
Figura 4. Durante il ciclo di Krebs, ogni piruvato generato dalla glicolisi viene convertito in una molecola di acetil CoA a due atomi di carbonio. L’acetil CoA viene sistematicamente elaborato attraverso il ciclo e produce molecole di NADH, FADH2 e ATP ad alta energia.
Guarda questa animazione per osservare il ciclo di Krebs.
La molecola di piruvato a tre atomi di carbonio generata durante la glicolisi si sposta dal citoplasma nella matrice mitocondriale, dove viene convertita dall’enzima piruvato deidrogenasi in una molecola di acetil coenzima A (acetil CoA) a due atomi di carbonio. Questa reazione è una reazione di decarbossilazione ossidativa. Converte il piruvato a tre atomi di carbonio in una molecola di acetil COA a due atomi di carbonio, rilasciando anidride carbonica e trasferendo due elettroni che si combinano con NAD+ per formare NADH. L’acetil CoA entra nel ciclo di Krebs combinandosi con una molecola a quattro atomi di carbonio, ossaloacetato, per formare il citrato di molecola a sei atomi di carbonio, o acido citrico, allo stesso tempo rilasciando la molecola di coenzima A.
La molecola di citrato a sei atomi di carbonio viene sistematicamente convertita in una molecola a cinque atomi di carbonio e quindi in una molecola a quattro atomi di carbonio, terminando con l’ossaloacetato, l’inizio del ciclo. Lungo la strada, ogni molecola di citrato produrrà un ATP, un FADH2 e tre NADH. Il FADH2 e il NADH entreranno nel sistema di fosforilazione ossidativa situato nella membrana mitocondriale interna. Inoltre, il ciclo di Krebs fornisce i materiali di partenza per elaborare e abbattere proteine e grassi.
Per iniziare il ciclo di Krebs, la citrato sintasi combina acetil CoA e ossaloacetato per formare una molecola di citrato a sei atomi di carbonio; Il CoA viene successivamente rilasciato e può combinarsi con un’altra molecola di piruvato per ricominciare il ciclo. L’enzima aconitasi converte il citrato in isocitrato. In due fasi successive di decarbossilazione ossidativa, due molecole di CO2 e due molecole di NADH vengono prodotte quando l’isocitrato deidrogenasi converte l’isocitrato nell’α-chetoglutarato a cinque atomi di carbonio, che viene quindi catalizzato e convertito nel succinil COA a quattro atomi di carbonio da α-chetoglutarato deidrogenasi. L’enzima succinil CoA deidrogenasi converte quindi succinil CoA in succinato e forma la molecola ad alta energia GTP, che trasferisce la sua energia all’ADP per produrre ATP. Succinato deidrogenasi poi converte succinato in fumarato, formando una molecola di FADH2. Fumarasi poi converte fumarato in malato, che malato deidrogenasi poi converte di nuovo in ossaloacetato riducendo NAD + a NADH. L’ossaloacetato è quindi pronto per essere combinato con il successivo acetil CoA per ricominciare il ciclo di Krebs (vedere Figura 4). Per ogni giro del ciclo, vengono creati tre NADH, un ATP (tramite GTP) e un FADH2. Ogni carbonio di piruvato viene convertito in CO2, che viene rilasciato come sottoprodotto della respirazione ossidativa (aerobica).
Fosforilazione ossidativa e catena di trasporto degli elettroni
La catena di trasporto degli elettroni (ETC) utilizza il NADH e il FADH2 prodotti dal ciclo di Krebs per generare ATP. Gli elettroni di NADH e FADH2 vengono trasferiti attraverso complessi proteici incorporati nella membrana mitocondriale interna da una serie di reazioni enzimatiche. La catena di trasporto degli elettroni consiste in una serie di quattro complessi enzimatici (Complesso I – Complesso IV) e due coenzimi (ubiquinone e citocromo c), che fungono da portatori di elettroni e pompe protoniche utilizzate per trasferire gli ioni H+ nello spazio tra le membrane mitocondriali interne ed esterne (Figura 5). L’ETC accoppia il trasferimento di elettroni tra un donatore (come NADH) e un accettore di elettroni (come O2) con il trasferimento di protoni (ioni H+) attraverso la membrana mitocondriale interna, consentendo il processo di fosforilazione ossidativa. In presenza di ossigeno, l’energia viene passata, gradualmente, attraverso i portatori di elettroni per raccogliere gradualmente l’energia necessaria per attaccare un fosfato all’ADP e produrre ATP. Il ruolo dell’ossigeno molecolare, O2, è come l’accettore terminale dell’elettrone per ECC. Ciò significa che una volta che gli elettroni sono passati attraverso l’intero ETC, devono essere passati ad un’altra molecola separata. Questi elettroni, O2 e ioni H + dalla matrice si combinano per formare nuove molecole d’acqua. Questa è la base per il tuo bisogno di respirare ossigeno. Senza ossigeno, il flusso di elettroni attraverso l’ETC cessa.
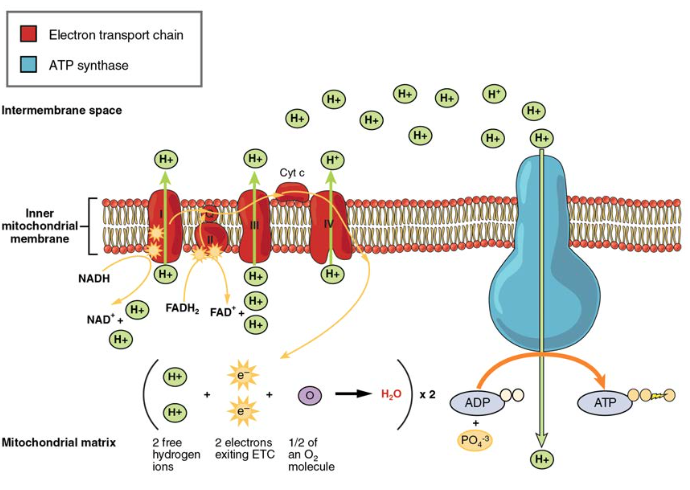
Figura 5. La catena di trasporto degli elettroni è una serie di portatori di elettroni e pompe ioniche che vengono utilizzate per pompare gli ioni H+ dalla matrice mitocondriale interna.
Guarda questo video per conoscere la catena di trasporto degli elettroni.
Gli elettroni rilasciati da NADH e FADH2 vengono passati lungo la catena da ciascuno dei portatori, che vengono ridotti quando ricevono l’elettrone e ossidati quando lo passano al vettore successivo. Ognuna di queste reazioni rilascia una piccola quantità
di energia, che viene utilizzata per pompare ioni H+ attraverso la membrana interna. L’accumulo di questi protoni nello spazio tra le membrane crea un gradiente protonico rispetto alla matrice mitocondriale.
Inoltre incastonato nella membrana mitocondriale interna è un complesso stupefacente del poro della proteina chiamato ATP sintasi. Efficacemente, è una turbina che è alimentata dal flusso degli ioni di H + attraverso la membrana interna giù un gradiente e nella matrice mitocondriale. Mentre gli ioni H + attraversano il complesso, l’albero del complesso ruota. Questa rotazione consente ad altre porzioni di ATP sintasi di incoraggiare ADP e Pi a creare ATP. Nel contabilizzare il numero totale di ATP prodotto per molecola di glucosio attraverso la respirazione aerobica, è importante ricordare i seguenti punti:
- Una rete di due ATP viene prodotta attraverso la glicolisi (quattro prodotti e due consumati durante la fase di consumo energetico). Tuttavia, questi due ATP sono utilizzati per trasportare il NADH prodotto durante la glicolisi dal citoplasma nei mitocondri. Pertanto, la produzione netta di ATP durante la glicolisi è zero.
- In tutte le fasi dopo la glicolisi, il numero di ATP, NADH e FADH2 prodotti deve essere moltiplicato per due per riflettere il modo in cui ogni molecola di glucosio produce due molecole di piruvato.
- Nell’ETC, vengono prodotti circa tre ATP per ogni NADH ossidato. Tuttavia, solo circa due ATP sono prodotti per ogni FADH2 ossidato. Gli elettroni di FADH2 producono meno ATP, perché iniziano in un punto inferiore dell’ETC (Complesso II) rispetto agli elettroni di NADH (Complesso I) (Figura 5).
Pertanto, per ogni molecola di glucosio che entra nella respirazione aerobica, vengono prodotti un totale netto di 36 ATPS (vedi Figura 6).
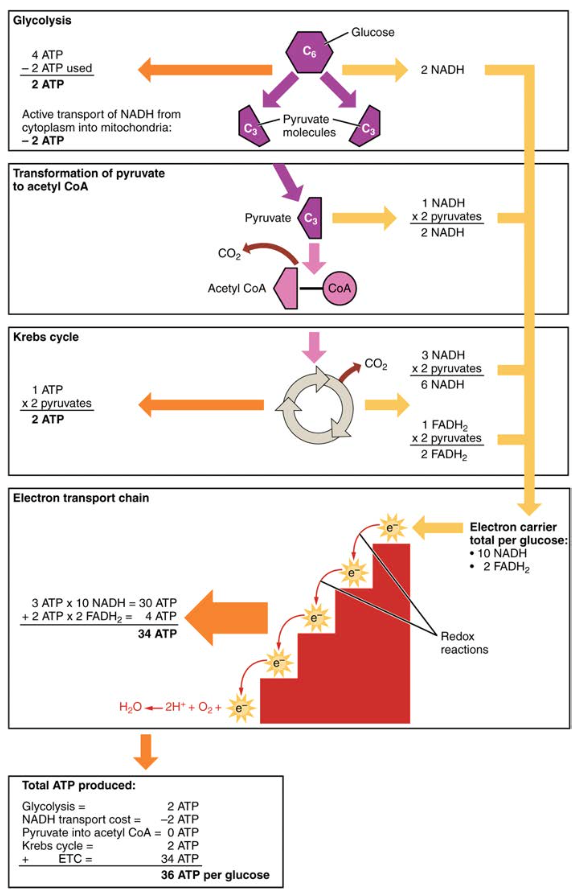
Figura 6. Il metabolismo dei carboidrati comporta la glicolisi, il ciclo di Krebs e la catena di trasporto degli elettroni.
Gluconeogenesi
La gluconeogenesi è la sintesi di nuove molecole di glucosio da piruvato, lattato, glicerolo o aminoacidi alanina o glutammina. Questo processo avviene principalmente nel fegato durante periodi di basso glucosio, cioè in condizioni di digiuno, fame e diete a basso contenuto di carboidrati. Quindi, la domanda può essere sollevata sul perché il corpo creerebbe qualcosa che ha appena speso una discreta quantità di sforzi per abbattere? Alcuni organi chiave, incluso il cervello, possono utilizzare solo il glucosio come fonte di energia; pertanto, è essenziale che il corpo mantenga una concentrazione minima di glucosio nel sangue. Quando la concentrazione di glucosio nel sangue scende al di sotto di quel certo punto, il nuovo glucosio viene sintetizzato dal fegato per aumentare la concentrazione nel sangue alla normalità.
La gluconeogenesi non è semplicemente il contrario della glicolisi. Ci sono alcune differenze importanti (Figura 7). Il piruvato è un materiale di partenza comune per la gluconeogenesi. Innanzitutto, il piruvato viene convertito in ossaloacetato. L’ossaloacetato funge quindi da substrato per l’enzima fosfoenolpiruvato carbossichinasi (PEPCK), che trasforma l’ossaloacetato in fosfoenolpiruvato (PEP). Da questa fase, la gluconeogenesi è quasi l’inverso della glicolisi. Il PEP viene riconvertito in 2-fosfoglicerato, che viene convertito in 3-fosfoglicerato. Quindi, il 3-fosfoglicerato viene convertito in 1,3 bisfosfoglicerato e quindi in gliceraldeide-3-fosfato. Due molecole di gliceraldeide-3-fosfato si combinano quindi per formare fruttosio-1-6-bisfosfato, che viene convertito in fruttosio 6-fosfato e quindi in glucosio-6-fosfato. Infine, una serie di reazioni genera glucosio stesso. Nella gluconeogenesi (rispetto alla glicolisi), l’enzima esochinasi viene sostituito da glucosio-6-fosfatasi e l’enzima fosfofruttochinasi-1 viene sostituito da fruttosio-1,6-bisfosfatasi. Questo aiuta la cellula a regolare la glicolisi e la gluconeogenesi indipendentemente l’una dall’altra.
Come sarà discusso come parte della lipolisi, i grassi possono essere suddivisi in glicerolo, che può essere fosforilato per formare fosfato diidrossiacetone o DHAP. DHAP può entrare nella via glicolitica o essere utilizzato dal fegato come substrato per la gluconeogenesi.
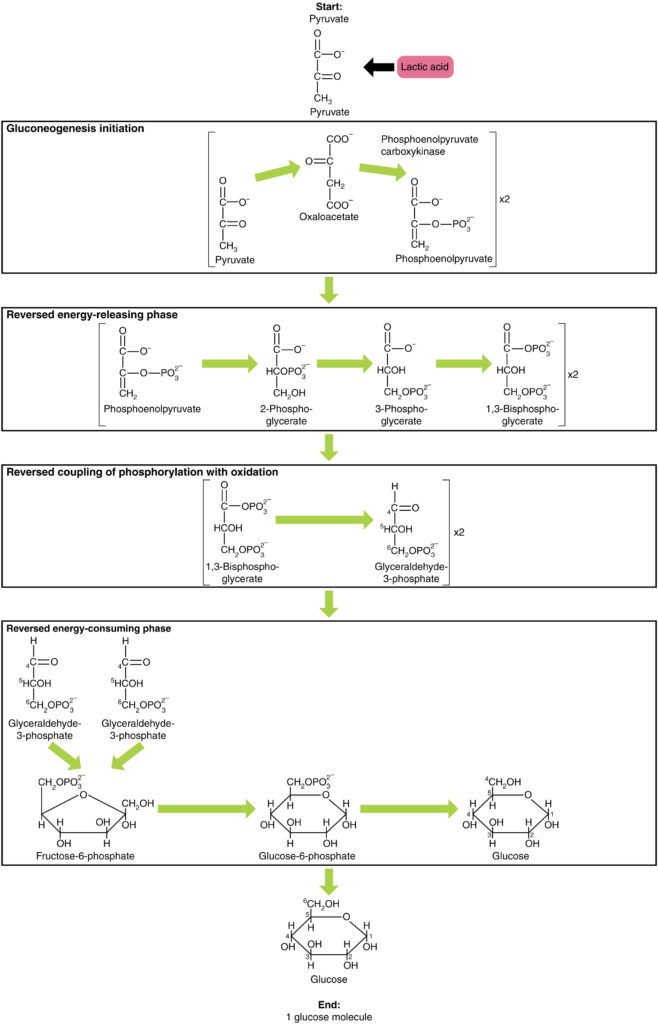
Figura 7. Fare clic per visualizzare un’immagine ingrandita. La gluconeogenesi è la sintesi del glucosio da piruvato, lattato, glicerolo, alanina o glutammato.
Invecchiamento e il tasso metabolico del corpo
Il tasso metabolico del corpo umano diminuisce quasi il 2 per cento per decennio dopo i 30 anni. I cambiamenti nella composizione corporea, compresa la ridotta massa muscolare magra, sono per lo più responsabili di questa diminuzione. La perdita più drammatica di massa muscolare, e conseguente declino del tasso metabolico, si verifica tra i 50 ei 70 anni di età. La perdita di massa muscolare è l’equivalente di una forza ridotta, che tende a inibire gli anziani dall’impegnarsi in un’attività fisica sufficiente. Ciò si traduce in un sistema di feedback positivo in cui la ridotta attività fisica porta a una perdita muscolare ancora maggiore, riducendo ulteriormente il metabolismo.
Ci sono diverse cose che possono essere fatte per aiutare a prevenire cali generali nel metabolismo e per combattere contro la natura ciclica di questi cali. Questi includono fare colazione, mangiare piccoli pasti frequentemente, consumare un sacco di proteine magre, bere acqua per rimanere idratati, esercitare (incluso l’allenamento della forza) e dormire a sufficienza. Queste misure possono aiutare a mantenere i livelli di energia da cadere e frenare la voglia di un aumento del consumo calorico da spuntini eccessivi. Mentre queste strategie non sono garantite per mantenere il metabolismo, aiutano a prevenire la perdita muscolare e possono aumentare i livelli di energia. Alcuni esperti suggeriscono anche di evitare lo zucchero, che può portare all’accumulo di grasso in eccesso. Cibi piccanti e tè verde potrebbero anche essere utili. Perché lo stress attiva il rilascio di cortisolo e il cortisolo rallenta il metabolismo, evitando lo stress, o almeno praticando tecniche di rilassamento, può anche aiutare.
Capitolo Recensione
Gli enzimi metabolici catalizzano le reazioni cataboliche che abbattono i carboidrati contenuti nel cibo. L’energia rilasciata viene utilizzata per alimentare le cellule e i sistemi che compongono il tuo corpo. L’energia in eccesso o non utilizzata viene immagazzinata come grasso o glicogeno per un uso successivo. Il metabolismo dei carboidrati inizia in bocca, dove l’enzima amilasi salivare inizia a scomporre gli zuccheri complessi in monosaccaridi. Questi possono poi essere trasportati attraverso la membrana intestinale nel flusso sanguigno e poi ai tessuti del corpo. Nelle cellule, il glucosio, uno zucchero a sei atomi di carbonio, viene elaborato attraverso una sequenza di reazioni in zuccheri più piccoli e l’energia immagazzinata all’interno della molecola viene rilasciata. Il primo passo del catabolismo dei carboidrati è la glicolisi, che produce piruvato, NADH e ATP. In condizioni anaerobiche, il piruvato può essere convertito in lattato per mantenere la glicolisi funzionante. In condizioni aerobiche, il piruvato entra nel ciclo di Krebs, chiamato anche ciclo dell’acido citrico o ciclo dell’acido tricarbossilico. Oltre all’ATP, il ciclo di Krebs produce molecole ad alta energia FADH2 e NADH, che forniscono elettroni al processo di fosforilazione ossidativa che genera più molecole di ATP ad alta energia. Per ogni molecola di glucosio che viene elaborata in glicolisi, una rete di 36 ATPS può essere creata dalla respirazione aerobica.
In condizioni anaerobiche, la produzione di ATP è limitata a quelle generate dalla glicolisi. Mentre un totale di quattro ATPS sono prodotti dalla glicolisi, due sono necessari per iniziare la glicolisi, quindi c’è una resa netta di due molecole di ATP.
In condizioni di basso glucosio, come digiuno, fame o diete a basso contenuto di carboidrati, il glucosio può essere sintetizzato da lattato, piruvato, glicerolo, alanina o glutammato. Questo processo, chiamato gluconeogenesi, è quasi il contrario della glicolisi e serve a creare molecole di glucosio per gli organi glucosio-dipendenti, come il cervello, quando i livelli di glucosio scendono al di sotto del normale.
Self Check
Rispondi alle domande qui sotto per vedere quanto bene comprendi gli argomenti trattati nella sezione precedente.
Domande di pensiero critico
- Spiegare come il glucosio viene metabolizzato per produrre ATP.
- Discutere il meccanismo cellule impiegano per creare un gradiente di concentrazione per garantire l’assorbimento continuo di glucosio dal flusso sanguigno.
Glossario
polisaccaridi: carboidrati complessi costituiti da molti monosaccaridi
monosaccaride: molecola di zucchero monomerica più piccola
amilasi salivare: enzima digestivo che si trova nella saliva e inizia la digestione dei carboidrati in bocca
la respirazione cellulare: la produzione di ATP da ossidazione del glucosio attraverso la glicolisi, ciclo di Krebs e la fosforilazione ossidativa
la glicolisi: serie di reazioni metaboliche che scinde il glucosio in piruvato e produce ATP
piruvato: tre emissioni di carbonio prodotto finale della glicolisi e del materiale di partenza, che viene convertito in acetil-CoA che entra
ciclo di Krebs: chiamato anche ciclo dell’acido citrico o ciclo degli acidi tricarbossilici, converte il piruvato in CO2 e ad alta energia FADH2, NADH, e molecole di ATP
ciclo dell’acido citrico o ciclo dell’acido tricarbossilico (TCA): chiamato anche il ciclo di Krebs o ciclo degli acidi tricarbossilici; converte il piruvato in CO2 e ad alta energia FADH2, NADH, e molecole di ATP
consumo energetico fase prima fase della glicolisi, in cui due molecole di ATP sono necessari per avviare la reazione
glucosio-6-fosfato: fosforilata glucosio prodotta nella prima fase della glicolisi
Esochinasi: enzima cellulare, trovato nella maggior parte dei tessuti, che converte il glucosio in glucosio-6-fosfato all’assorbimento nella cella
la Glucochinasi: cellularenzyme, trovato nel fegato, che converte il glucosio in glucosio-6-fosfato all’assorbimento nella cella
ener getico fase: seconda fase della glicolisi, durante la quale l’energia è prodotta
terminale accettore di elettroni: La produzione di ATP percorso in cui gli elettroni sono passati attraverso una serie di reazioni di ossido-riduzione che si forma acqua e produce un gradiente protonico
ciclo di Krebs: chiamato anche ciclo dell’acido citrico o ciclo degli acidi tricarbossilici, converte il piruvato in CO2 e ad alta energia FADH2, NADH, e molecole di ATP
la catena di trasporto degli elettroni (ETC): la produzione di ATP percorso in cui gli elettroni sono passati attraverso una serie di reazioni di ossido-riduzione che si forma acqua e produce un gradiente protonico
la fosforilazione ossidativa: processo che converte ad alta energia NADH e FADH2 in ATP
ATP sintasi, proteine: complesso del poro che crea ATP
la Gluconeogenesi: processo di sintesi di glucosio da piruvato o altre molecole