Introduzione
L’interazione riuscita con il nostro ambiente sensoriale richiede un intricato equilibrio di due meccanismi di selezione attenzionale—quello top-down e bottom-up. Dirigendosi verso il corridoio dei prodotti del tuo supermercato locale con l’obiettivo di raccogliere pochi ingredienti necessari per l’insalata di mango, si impegna la distribuzione di un sistema di attenzione mirato o dall’alto verso il basso in modo tale da cercare attivamente tutti gli ingredienti necessari tra la moltitudine di scelte di prodotti. Tuttavia, se si sente una suoneria di un telefono cellulare, molto probabilmente catturare la vostra attenzione e interrompere la ricerca. Tale interruzione si verifica in un bottom-up, o stimolo-driven, moda per cui una mera salienza dello stimolo, il fatto che l’anello è diverso da altri suoni nel vostro ambiente, lo ritiene degno di selezione. Lo scenario descritto sottolinea l’importanza della selezione orientata agli obiettivi e guidata dagli stimoli per il comportamento e indica un sottile equilibrio che deve esistere tra i due sistemi di attenzione per prevenire la “visione a tunnel” da un lato e la completa incapacità di concentrarsi sull’altro.
Selezione dall’alto verso il basso e dal basso verso l’alto: Comportamento
Diversi decenni di ricerca comportamentale sono stati dedicati a dimostrare che la distribuzione dell’attenzione può essere controllata dalle intenzioni dell’osservatore e dalla salienza dello stimolo fisico. Gran parte delle prove comportamentali per l’allocazione dell’attenzione dall’alto verso il basso e dal basso verso l’alto è stata ampiamente rivista altrove (Johnston e Dark, 1986; Egeth e Yantis, 1997). Per riassumere, gli studi che dimostrano gli effetti del controllo dell’attenzione dall’alto verso il basso mostrano che l’attenzione può essere assegnata con successo a posizioni spaziali, caratteristiche, oggetti, ecc., a seguito della presenza di segnali esogeni o endogeni (Eriksen e Hoffman, 1972; Posner, 1980; Posner et al., 1980), o aspettative stabilite da conoscenze precedenti o da contingenze dello stimolo (Shaw, 1978; Moore e Egeth, 1998; Geng e Behrmann, 2002, 2005; Shomstein e Yantis, 2004a; Drummond e Shomstein, 2010). Le prove a sostegno dell’allocazione attentiva dal basso verso l’alto si sono basate su vari paradigmi di cattura dell’attenzione, in cui i partecipanti sono impegnati in una ricerca dall’alto verso il basso e la loro attenzione è deviata verso gli stimoli irrilevanti del compito, dimostrando che l’attenzione è catturata dai singoli personaggi (oggetto unico; Yantis e Jonides, 1990; Theeuwes, 1991; Folk et al., 2002) e onset bruschi (Yantis e Jonides, 1984; Theeuwes, 1991; Koshino et al., 1992; Juola et al., 1995).
Mentre la maggior parte dei primi studi si concentrava sulla dimostrazione di prove per la selezione attentiva dall’alto verso il basso e dal basso verso l’alto, gli studi più recenti hanno spostato la loro attenzione sull’esame del modo in cui i due sistemi di selezione attentiva interagiscono. Questa linea di indagine è alimentata dalle osservazioni che, al fine di selezionare efficacemente le informazioni rilevanti per l’attività (ad esempio, gli ingredienti per l’insalata), è necessario inibire attivamente le informazioni irrilevanti per l’attività che altrimenti distoglierebbero l’attenzione dall’attività in questione. Il rovescio della medaglia di questa logica è che meno si concentra sulle informazioni relative alle attività, più ne conseguirà la cattura. È stato dimostrato sperimentalmente che lo stato di attenzione dell’osservatore predice quale tipo di informazione, e in che misura, alla fine catturerà l’attenzione (Folk et al., 1992, 2002; Bacon e Egeth, 1994; Gibson e Kelsey, 1998). Ad esempio, Folk et al. (2002) ha dimostrato che quando si cerca una lettera rossa, un osservatore sarà più facilmente catturato da uno stimolo irrilevante nella periferia se tale stimolo è rosso o corrisponde in qualche modo al modello di destinazione. Poiché le impostazioni di controllo dall’alto verso il basso dell’osservatore sono impostate per cercare una caratteristica rossa, è probabile che qualsiasi stimolo rosso catturi l’attenzione e potenzialmente interferisca con il controllo dall’alto verso il basso. Pertanto, con un compito di cattura, le strategie di ricerca attentiva possono essere distinte l’una dall’altra variando i livelli di somiglianza tra le proprietà di stimolo del bersaglio e dei distrattori. Più il bersaglio è simile al distrattore, più è difficile per l’osservatore evitare la cattura.
Il ruolo del lobo parietale nella selezione Top-Down e Bottom-Up: Neuroimaging
Varie tecniche di neuroimaging hanno fornito una forte evidenza per il coinvolgimento della corteccia parietale nell’orientamento top-down e bottom-up, con le prove ampiamente riviste altrove (Corbetta e Shulman, 2002, 2011; Behrmann et al., 2004). È stato dimostrato che le aree più comunemente attivate in seguito a segnali dall’alto verso il basso per partecipare a particolari posizioni, caratteristiche o oggetti si trovano lungo le parti dorsali della corteccia parietale. Tali aree includono il lobulo parietale inferiore (IPL), le regioni dorsomediali indicate come lobulo parietale superiore (SPL), così come le regioni più mediali lungo il giro precuneo (Yantis et al., 2002; Giesbrecht et al., 2003; Liu et al., 2003; Yantis e Serences, 2003; Figura 1). Diversi compiti dall’alto verso il basso sono stati dimostrati per coinvolgere con successo le regioni dorsali della corteccia parietale, vale a dire quelle che coinvolgono lo spazio (Kastner et al., 1999; Corbetta et al., 2000; Hopfinger et al., 2000; Shomstein e Behrmann, 2006; Chiu e Yantis, 2009; Greenberg et al., 2010) così come spostamenti non spaziali di attenzione (Giesbrecht et al., 2003; Yantis e Serences, 2003; Shomstein e Yantis, 2004b, 2006; Tamber-Rosenau et al., 2011).
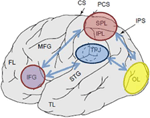
Figura 1. Rappresentazione schematica di importanti punti di riferimento anatomici proiettati sulla superficie laterale del cervello umano. Il lobulo parietale superiore (SPL) ed il lobulo parietale inferiore (IPL) sono regioni all’interno della parte dorsale della corteccia parietale che sottoservisce l’orientamento attenzionale dall’alto verso il basso. La giunzione temporo-parietale (TPJ) è una regione all’interno della corteccia parietale ventrale che sottomette l’orientamento attenzionale dal basso verso l’alto. Entrambi, SPL e TPJ, sono pensati per suscitare segnali di controllo responsabili delle successive modulazioni attenzionali osservate sulle regioni sensoriali, in questo caso modulando (etichettate con frecce blu scuro) l’attività visivamente evocata nel lobo occipitale (OL). Inoltre, le aree lungo il giro frontale inferiore (IFG) e la giunzione frontale inferiore (IFJ) sono pensate per servire come aree di convergenza per il controllo attenzionale guidato dallo stimolo e dall’alto verso il basso (contrassegnate da frecce bidirezionali blu chiaro).
In un tipico compito volto a coinvolgere l’allocazione dell’attenzione dall’alto verso il basso, agli individui vengono mostrati due flussi di presentazione visiva seriale rapida (RSVP) posizionati perifericamente e vengono inizialmente istruiti a monitorare un flusso per un cue (ad esempio, una cifra tra il flusso di lettere). L’identità della stecca indica se il soggetto deve mantenere l’attenzione sul flusso corrente o spostare l’attenzione sull’altro flusso (Yantis et al., 2002; Yantis e Serences, 2003). Due risultati principali sono osservati in tali paradigmi. Il primo ha a che fare con una maggiore attivazione all’interno delle regioni sensoriali che rappresentano la posizione frequentata al momento (ad esempio, una maggiore attività all’interno delle regioni visive primarie di sinistra quando viene frequentato il flusso RSVP destro). Questa scoperta fornisce prove concrete che i partecipanti stanno frequentando un luogo specifico e che l’attenzione modula la forza della risposta sensoriale (vedi Figura 1; Moran e Desimone, 1985; O’Craven et al., 1997). La seconda scoperta ha a che fare con l’osservazione che le regioni dorsali del lobo parietale sono selettivamente attivate da spostamenti di attenzione dall’alto verso il basso. Si osserva che il tempo di attività SPL/IPL è di natura transitoria, suggerendo che quest’area della corteccia parietale è la fonte di un breve segnale di controllo attentivo per spostare gli stati attenti in modo top-down (Yantis et al., 2002).
Diversi studi di fMRI hanno documentato che la cattura attentiva bottom-up, mediata dalla salienza e / o rilevanza dello stimolo, è sottomessa dalla giunzione temporo-parietale (TPJ; Figura 1). Ad esempio, quando i soggetti assistono e monitorano un cambiamento in uno stimolo visivo o uditivo, presentato simultaneamente, l’attivazione delle regioni TPJ del lobo parietale è migliorata. Oltre all’apparente sensibilità agli stimoli rilevanti, il TPJ viene attivato anche in risposta a eventi potenzialmente nuovi (inaspettati o poco frequenti) quando un organismo è impegnato in un contesto comportamentale neutro o quando è impegnato in un compito (Marois et al., 2000; Downar et al., 2002; Serences et al., 2005; Corbetta et al., 2008; Asplund et al., 2010; Diquattro e Geng, 2011; Geng e Mangun, 2011). Questa attivazione avviene indipendentemente dalla modalità (uditiva, tattile e visiva) in cui viene consegnato l’input, riflettendo la natura multisensoriale di TPJ (ma vedi Downar et al., 2001).
In un tipico compito che esamina il meccanismo neurale della cattura attentiva bottom-up, ai partecipanti viene presentato un flusso RSVP di elementi al centro del display e viene chiesto di identificare un target predefinito (ad esempio, identificare la lettera rossa presentata all’interno di un flusso RSVP di non target bianchi). Una parte delle prove contiene un distrattore saliente irrilevante per attività presentato a vari intervalli di tempo prima dell’inizio del bersaglio, mentre altre prove contengono solo il distrattore saliente (cioè, senza il bersaglio). Le prove “Target-distractor” sono utilizzate per valutare l’entità della cattura, dimostrando che il distrattore irrilevante per il compito è in realtà saliente, producendo così una diminuzione della precisione del bersaglio. Le prove “distractor-in-isolation” sono utilizzate per ulteriori analisi poiché tali prove consentono l’esame dell’attività suscitata dal distrattore saliente senza contaminazione dai processi correlati all’obiettivo. Diversi risultati importanti emergono da tali paradigmi. In primo luogo, quando i distrattori sono spazialmente separati dalla posizione di destinazione, i distrattori di cattura sono accompagnati da una maggiore attività corticale nelle regioni corrispondenti della corteccia sensoriale (ad esempio, corteccia visiva organizzata retinotopicamente; vedere Figura 1). Tali risultati forniscono una forte evidenza che durante la cattura, l’attenzione spaziale viene effettivamente catturata nella posizione spaziale occupata dal distrattore (Serences et al., 2005). In secondo luogo, le regioni ventrali della corteccia parietale, principalmente all’interno del TPJ, sono selettivamente attivate da spostamenti di attenzione dal basso verso l’alto, involontari. Proprio come l’attività all’interno dell’SPL per l’orientamento dall’alto verso il basso, il corso temporale dell’attività osservato su TPJ è di natura transitoria, suggerendo che questa regione è la fonte di un breve segnale di controllo dell’attenzione per spostare l’attenzione in modo bottom-up.
Va notato che mentre questa revisione è focalizzata sull’affrontare le funzioni cognitive della corteccia parietale posteriore, altre regioni, in particolare quelle all’interno della corteccia frontale sono anche reclutate per l’allocazione dell’attenzione dall’alto verso il basso e dal basso verso l’alto. Tali regioni includono la corteccia frontale ventrale (VFC), i campi dell’occhio frontale (FEF), la giunzione frontale inferiore (IFJ) e il giro frontale inferiore (IFG; Corbetta e Shulman, 2002, 2011; Serences et al., 2005; Asplund et al., 2010; Diquattro e Geng, 2011).
Il ruolo del lobo parietale nella selezione Top-Down e Bottom-Up: Neuropsicologia
Storicamente i ricercatori si sono basati criticamente su studi neuropsicologici di pazienti con negligenza emispaziale (un disturbo dell’allocazione spaziale dell’attenzione all’emispazio sinistro) per ottenere informazioni sulle funzioni cognitive associate al lobo parietale. Nella letteratura neuropsicologica classica, la corteccia parietale, nella sua interezza, era generalmente considerata il sito della lesione primaria per la negligenza emispaziale. Questo punto di vista, elaborato in dettaglio dai primi ricercatori (Critchley, 1953; McFire e Zangwill, 1960; Piercy, 1964) ha chiaramente riconosciuto l’associazione tra la lesione parietale e la conseguente negligenza. Questa prospettiva è stata in gran parte mantenuta attraverso gli 1980 quando Posner e colleghi (1984) hanno usato il paradigma cueing visuospaziale nascosto per mostrare che il danno al lobo parietale produce un deficit nell’operazione di “disimpegno” (retrazione dell’attenzione da una posizione e spostamento a un’altra) quando il bersaglio è controlaterale alla lesione. Tuttavia, nonostante questo importante progresso nella comprensione della base neurale dell’attenzione e in particolare del ruolo di “disimpegno” della corteccia parietale, i loro risultati assumono un singolo sito corticale (corteccia parietale) e una singola capacità funzionale (“disimpegno”). In contrasto con questo approccio più monolitico al cervello (corteccia parietale) e al comportamento (disimpegno attenzionale), il recente lavoro comportamentale e neuroimaging (esaminato sopra e altrove) suggerisce che sia la regione corticale che il comportamento attenzionale associato possono essere suddivisi in profili qualitativamente diversi.
Dato che la segregazione delle reti corticali nella top-down e bottom-up di processi, un evidente previsione è che i danni al superiore porzioni del lobulo parietale (sussumere SPL) dovrebbe produrre un deficit di goal-directed attenzione, orientamento, considerando che il danno per le porzioni inferiori del lobulo parietale (sussumere TPJ) comporterebbe un deficit associati con stimolo catturare l’attenzione. Nella misura in cui queste corrispondenze cervello-comportamento sono stati esplorati nella letteratura neuropsicologica, questa previsione non è ovviamente confermata. Ad esempio, i sintomi clinici di negligenza emispaziale sono fortemente associati a danni alle porzioni inferiori del lobo parietale, che include TPJ, piuttosto che a porzioni superiori come SPL (Friedrich et al., 1998; Shomstein et al., 2010; Corbetta e Shulman, 2011). Questo è in qualche modo in contrasto con la letteratura neuroimaging, che suggerisce che il ruolo di TPJ è nella cattura dell’attenzione, piuttosto che nell’orientamento volontario dell’attenzione, il dominio in cui i pazienti trascurati sembrano avere più difficoltà. A complicare ulteriormente le cose, è stato notato che le lesioni che coinvolgono esclusivamente SPL, solo raramente producono prove cliniche di negligenza (Vallar e Perani, 1986). Un altro recente studio con pazienti con lesioni centrato principalmente su TPJ e STG ma conservato SPL, Corbetta et al. (2005) ha dimostrato che la negligenza spaziale, così come il suo recupero, era associata al ripristino dell’attività sia nelle regioni ventrale temporo-parietale che dorsale parietale (vedi Corbetta e Shulman, 2011 per una revisione). Mentre interessante ed emozionante nelle sue conclusioni, questo ultimo studio non differenzia il contributo relativo delle vie dorsali e ventrali a diversi tipi di attenzione, dal momento che i pazienti sono stati testati solo su una variante del compito di cuing dell’attenzione spaziale nascosta di Posner, compito che si pensa di coinvolgere sia l’orientamento dell’attenzione dall’alto verso il basso che
Per distinguere tra il controllo dell’attenzione guidato dall’obiettivo e la cattura dell’attenzione saliente e per esaminare la loro mappatura rispettivamente su SPL e TPJ, uno studio recente ha adottato due paradigmi comportamentali, ciascuno mirato a una di queste forme di attenzione (Shomstein et al., 2010). Per esaminare l’integrità dell’orientamento attentivo top-down nei pazienti, è stato utilizzato un compito top-down che richiedeva ai partecipanti di spostare l’attenzione spaziale tra i flussi RSVP spazialmente separati (un compito che è stato utilizzato con successo per dimostrare l’attivazione SPL negli studi fMRI (Yantis et al., 2002)). Allo stesso modo, al fine di esaminare le capacità di orientamento attento bottom-up dei pazienti, una variante di Folk et al. (2002) è stato impiegato il paradigma di cattura contingente in cui i partecipanti hanno rilevato obiettivi che apparivano alla fissazione mentre i singleton di colore irrilevanti per attività venivano lampeggiati nella periferia. La misura in cui i distrattori irrilevanti interferiscono con il compito di rilevamento centrale è stata quindi utilizzata come misura della cattura attenzionale dal basso verso l’alto (Bacon e Egeth, 1994; Folk et al., 2002).
Le previsioni erano le seguenti: i pazienti con lesioni a porzioni superiori del lobo parietale (che interessano SPL) devono essere compromesse nell’attività di orientamento dell’attenzione dall’alto verso il basso (con prestazioni preservate nell’attività di cattura) mentre i pazienti con lesioni alle porzioni inferiori del lobo parietale (che interessano TPJ) devono essere compromesse nell’attività di cattura (con prestazioni risparmiate nell’attività dall’alto verso il basso). Una doppia dissociazione di questa forma non solo attesta le componenti indipendenti dell’attenzione, ma suggerisce anche che tali componenti attentive sono mediate da meccanismi neurali indipendenti. Otto pazienti con negligenza visuo-spaziale sono stati reclutati per lo studio e hanno completato due compiti, toccando l’orientamento attenzionale guidato dallo stimolo o diretto all’obiettivo. In base al loro profilo comportamentale, i pazienti sono stati ordinati in gruppi e la loro sovrapposizione di lesioni è stata esplorata (Figura 2A). I pazienti che hanno mostrato difficoltà con l’orientamento attentivo diretto all’obiettivo, come quantificato dall’indice di attenzione dall’alto verso il basso (Figura 2B), hanno presentato una sovrapposizione della lesione centrata su porzioni superiori del lobulo parietale (SPL subsumante) con lobulo parietale inferiore risparmiato (TPJ). I pazienti con lesione si sovrappongono centrati sulle porzioni inferiori del lobulo parietale (subsuming TPJ) ma risparmiati SPL eseguita normalmente sul compito di orientamento obiettivo-diretto, pur rimanendo immune alla cattura attenzionale (Figura 2C). I risultati di questo studio suggeriscono chiaramente che SPL e TPJ sono regioni anatomiche che sono necessariamente reclutate ai fini dell’orientamento top-down e bottom-up e che il danno a SPL e TPJ porta a disturbi dell’orientamento top-down e bottom-up rispettivamente.
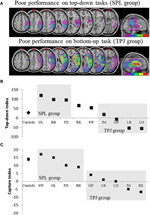
Figura 2. Risultati dello studio neuropsicologico volto a indagare il contributo relativo di SPL e TPJ all’orientamento top-down e bottom-up. (A) La lesione si sovrappone (sovrapposizione minima viola; sovrapposizione massima rossa) per i pazienti raggruppati per deficit comportamentali nell’orientamento attenzionale dall’alto verso il basso, etichettato il gruppo SPL (pannello superiore); e i pazienti raggruppati per deficit comportamentali nell’orientamento bottom-up, etichettato il gruppo TPJ (pannello inferiore). (B) Prestazioni comportamentali sull’attività top-down riassunte con un “Indice top-down” che quantifica le differenze tra gli spostamenti spaziali top-down effettuati da sinistra a destra e viceversa. I controlli e il gruppo lesionato TPJ mostrano efficienze simili nell’esecuzione di spostamenti spaziali, mentre i pazienti con lesioni SPL mostrano una diminuzione dell’efficienza. Il controllo di gruppo e i dati individuali del paziente (etichettati con le iniziali del paziente) sono tracciati sull’ascissa. (C)” Capture index ” è una misura di attenzione bottom-up e quantifica la misura in cui distrattori compito irrilevante catturare l’attenzione lontano dal compito. I controlli e il gruppo lesionato SPL mostrano valori di acquisizione simili, in modo che entrambi i gruppi vengano acquisiti dai distrattori irrilevanti dell’attività. Il gruppo lesionato TPJ mostra un indice di cattura molto ridotto (mancata acquisizione). Si noti che i pazienti sono stati inseriti nel gruppo SPL o TPJ in base al comportamento, piuttosto che in base alla lesione, quindi si noti la coerenza con cui i pazienti finiscono nel gruppo corrispondente.
Interazione tra selezione top-Down e Bottom-Up
Sebbene apparentemente vi sia una forte associazione tra orientamento diretto all’obiettivo e SPL e orientamento guidato dallo stimolo e TPJ, dati di Shomstein et al. (2010) lo studio del paziente suggerisce che questi due sistemi non sono del tutto indipendenti. Questa conclusione è supportata dalla constatazione che i pazienti con danno SPL esibivano un modello di prestazioni etichettato come ” hyper capture.”A differenza dei controlli, per i quali solo il distrattore colorato target ha catturato l’attenzione (portando a una minore precisione del bersaglio), anche i distrattori colorati irrilevanti si sono rivelati fonte di distrazione per i pazienti con lesione SPL. Inoltre, mentre per i controlli l’attenzione è stata catturata dai distrattori solo quando hanno preceduto l’inizio del bersaglio, per i pazienti con lesioni SPL l’attenzione è stata catturata anche dai distrattori presentati contemporaneamente al bersaglio. Questo modello di prestazioni può essere spiegato dal seguente framework: SPL è responsabile della guida top-down dell’attenzione che include la determinazione degli aspetti degli stimoli che sono rilevanti per il compito (ad esempio, ricerca di target rosso; Corbetta e Shulman, 2002; Serences et al., 2005). Questo set di attenzione vincola quindi TPJ, in modo tale che il meccanismo di cattura dell’attenzione mediato da TPJ sia attivato solo dalle informazioni rilevanti dell’attività (ad esempio, distrattori rossi che catturano l’attenzione e distrattori grigi che non catturano l’attenzione durante la ricerca di un bersaglio rosso). L’assenza di SPL impedisce l’istituzione di un set di attenzione pertinente al compito e quindi qualsiasi stimolo, compito rilevante o meno, è ritenuto importante quindi catturare l’attenzione (ad esempio, distrattore irrilevante del compito che cattura l’attenzione per il gruppo SPL) indiscriminatamente.
È stato suggerito che SPL e TPJ potrebbero interagire in almeno uno dei due modi possibili. La prima possibilità è che TPJ funga da sistema di allarme che rileva stimoli comportamentalmente rilevanti ma manca dell’alta risoluzione spaziale, quindi quando viene rilevato uno stimolo comportamentalmente rilevante la sua posizione precisa viene fornita dallo SPL che memorizza le mappe spaziali (Kastner et al., 1999; Wojciulik e Kanwisher, 1999; Bisley e Goldberg, 2003; Silver et al., 2005). Una possibilità ipotetica correlata è che il meccanismo di cattura (che include TPJ) agisca come un interruttore di attività cognitiva in corso quando viene presentato uno stimolo comportamentale rilevante (Corbetta e Shulman, 2002, 2011). Il modello di attività “hyper-capture” osservato in pazienti con TPJ conservato ma SPL lesionato fornisce ulteriori prove per l’ipotesi che vede TPJ come emissione di un segnale di controllo che termina il compito a portata di mano fungendo così da interruttore (Corbetta e Shulman, 2002; Serences et al., 2005). Altri recenti studi di neuroimaging che impiegano vari paradigmi hanno fornito ulteriori prove per una relazione interattiva tra l’orientamento attentivo top-down e bottom-up, e successivamente per la relazione tra SPL e TPJ (Serences et al., 2005; Asplund et al., 2010; Diquattro e Geng, 2011).
Mentre l’evidenza di un’interazione tra i due sistemi di attenzione e i due substrati di attenzione (SPL e TPJ) è forte, ciò che rimane poco chiaro è se questa interazione sia diretta tra SPL e TPJ o se sia realizzata attraverso altre regioni intermedie. Come accennato in precedenza, le reti di orientamento attentivo top-down e bottom-up coinvolgono varie regioni all’interno della corteccia frontale, quindi è ragionevole ipotizzare che la convergenza tra i due sistemi possa essere compiuta tramite il lobo frontale. Due recenti studi che indagano l’interazione tra la selezione attentiva dall’alto verso il basso e dal basso verso l’alto hanno fornito prove per l’IFJ e l’IFG come possibili siti di convergenza tra la selezione guidata dallo stimolo e quella diretta all’obiettivo (Asplund et al., 2010; Diquattro e Geng, 2011). L’IFJ e l’IFG sembrano essere candidati ideali per tale interazione dato il loro coinvolgimento generale nell’attenzione e nel controllo cognitivo, nonché il suo coinvolgimento nella selezione spaziale e non spaziale (Koechlin et al., 2003; Brass et al., 2005).
Il ruolo del lobo parietale nella selezione Top-Down e Bottom-Up: Fisiologia
Mentre l’enfasi di questa recensione è stata posta prevalentemente sugli studi umani, una grande ricchezza di conoscenze sul coinvolgimento della corteccia parietale nell’orientamento dell’attenzione è stata raccolta dalle indagini sulla fisiologia delle scimmie (vedi recente recensione di Bisley e Goldberg, 2010). Tuttavia, quando si tratta di esaminare i contributi relativi di diverse regioni all’interno della corteccia parietale all’orientamento attentivo dall’alto verso il basso e dal basso verso l’alto, la letteratura sulla fisiologia delle scimmie non è all’altezza. La ragione principale di ciò è che all’interno della corteccia della scimmia non sembra esserci prova della stessa segregazione del controllo dall’alto verso il basso e dal basso verso l’alto. Invece, area leistungskurs laterale (LABBRO), originariamente pensato per essere coinvolti nella saccade di pianificazione (Gnadt e Andersen, 1988) è coinvolto nell’attenzione visiva e agisce come una priorità mappa in cui gli stimoli esterni sono rappresentati secondo la loro comportamentali priorità derivati in top-down o bottom-up il modo (Colby e Goldberg, 1999; Bisley e Goldberg, 2003, 2010; Balan e Gottlieb, 2006; Ipata et al., 2006; Buschman e Miller, 2007; Gottlieb e Balan, 2010).
Conclusione
Sebbene sulla corteccia parietale umana si conosca molto meno di quella della corteccia monkey omologa, studi recenti, che impiegano metodi neuroimaging e neuropsicologici, hanno iniziato a chiarire distinzioni funzionali e strutturali sempre più a grana fine. Questa recensione si è concentrata su recenti studi neuroimaging e neuropsicologici che chiariscono i ruoli cognitivi delle regioni dorsali e ventrali della corteccia parietale nell’orientamento attentivo dall’alto verso il basso e dal basso verso l’alto e sull’interazione tra i due meccanismi di allocazione attenzionale.
Dichiarazione sul conflitto di interessi
L’autore dichiara che la ricerca è stata condotta in assenza di rapporti commerciali o finanziari che potrebbero essere interpretati come un potenziale conflitto di interessi.
Riconoscimenti
Questo lavoro è stato sostenuto dal National Institutes of Health grant EY021644.
Asplund, CL, Todd, JJ, Snyder, AP, e Marois, R. (2010). Un ruolo centrale per la corteccia prefrontale laterale nell’attenzione mirata e stimolata. NAT. Neurosci. 13, 507–512.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Bacon, W. F., and Egeth, H. E. (1994). Overriding stimulus-driven attentional capture. Percetto. Psicofisi. 55, 485–496.
Pubmed Abstract/Pubmed Full Text
Balan, P. F., e Gottlieb, J. (2006). Integrazione di input esogeni in una mappa dinamica della salienza rivelata dall’attenzione perturbante. J. Neurosci. 26, 9239–9249.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Behrmann, M., Geng, JJ, and Shomstein, S. (2004). Corteccia parietale e attenzione. Curr. Opin. Neurobiolo. 14, 212–217.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Bisley, JW, and Goldberg, M. E. (2003). Attività neuronale nell’area intraparietale laterale e attenzione spaziale. Scienza 299, 81-86.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Bisley, JW, and Goldberg, M. E. (2010). Attenzione, intenzione e priorità nel lobo parietale. Annu. Rev. Neurosci. 33, 1–21.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Brass, M., Derrfuss, J., Forstmann, B. e von Cramon, D. Y. (2005). Il ruolo dell’area di giunzione frontale inferiore nel controllo cognitivo. Tendenze Cogn. Sic. 9, 314–316.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Buschman, TJ e Miller, EK (2007). Controllo dall’alto verso il basso e dal basso verso l’alto dell’attenzione nelle cortecce parietali prefrontali e posteriori. Scienza 315, 1860-1862.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Chiu, Y. C., and Yantis, S. (2009). Una fonte indipendente dal dominio di controllo cognitivo per i set di attività: spostando l’attenzione spaziale e cambiando le regole di categorizzazione. J. Neurosci. 29, 3930–3938.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Colby, C. L., and Goldberg, M. E. (1999). Spazio e attenzione nella corteccia parietale. Annu. Rev. Neurosci. 22, 319–349.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., Kincade, JM, Oinger, JM, McAvoy, MP, e Shulman, GL (2000). L’orientamento volontario è dissociato dal rilevamento del bersaglio nella corteccia parietale posteriore umana. NAT. Neurosci. 3, 292–297.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., Kincade, M. J., Lewis, C., Snyder, A. Z. e Sapir, A. (2005). Basi neurali e recupero dei deficit di attenzione spaziale nell’abbandono spaziale. NAT. Neurosci. 8, 1603–1610.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., Patel, G., and Shulman, G. L. (2008). Il sistema di riorientamento del cervello umano: dall’ambiente alla teoria della mente. Neurone 58, 306-324.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., and Shulman, G. L. (2002). Controllo dell’attenzione mirata e stimolata nel cervello. NAT. Rev. Neurosci. 3, 201–215.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., and Shulman, G. L. (2011). Trascuratezza spaziale e reti di attenzione. Annu. Rev. Neurosci. 34, 569–599.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Critchley, M. (1953). I lobi parietali. Londra, Regno Unito: Hafner Press.
Pubmed Abstract/Pubmed Full Text
Diquattro, N. E., and Geng, JJ (2011). La conoscenza contestuale configura le reti di controllo dell’attenzione. J. Neurosci. 31, 18026–18035.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Downar, J., Crawley, AP, Mikulis, DJ, and Davis, KD (2001). L’effetto della rilevanza del compito sulla risposta corticale ai cambiamenti negli stimoli visivi e uditivi: uno studio fMRI correlato agli eventi. Neuroimage 14, 1256-1267.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Downar, J., Crawley, AP, Mikulis, DJ, and Davis, KD (2002). Una rete corticale sensibile alla salienza dello stimolo in un contesto comportamentale neutro attraverso molteplici modalità sensoriali. J. Neurofisiolo. 87, 615–620.
Pubmed Abstract/Pubmed Full Text
Drummond, L., and Shomstein, S. (2010). Attenzione basata sugli oggetti: spostamento o incertezza? Attenzione. Percetto. Psicofisi. 72, 1743–1755.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Egeth, H. E., and Yantis, S. (1997). Attenzione visiva: controllo, rappresentazione e corso del tempo. Annu. Rev. Psicol. 48, 269–297.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Eriksen, CW e Hoffman, JE (1972). Caratteristiche temporali e spaziali della codifica selettiva da display visivi. Percetto. Psicofisi. 12, 201–204.
Folk, C. L., Leber, A. B., e Egeth, H. E. (2002). Ti ha fatto battere le palpebre! La cattura dell’attenzione contingente produce un battito di ciglia spaziale. Percetto. Psicofisi. 64, 741–753.
Pubmed Abstract/Pubmed Full Text
Folk, C. L., Remington, R. W., e Johnston, J. C. (1992). L’orientamento involontario sotto copertura dipende dalle impostazioni di controllo dell’attenzione. J. Scad. Psicol. Ronzio. Percetto. Eseguire. 18, 1030–1044.
Pubmed Abstract/Pubmed Full Text
Friedrich, FJ, Egly, R., Rafal, R. D. e Beck, D. (1998). Deficit di attenzione spaziale nell’uomo: un confronto tra lesioni della giunzione parietale e temporale-parietale superiore. Neuropsicologia 12, 193-207.
Pubmed Abstract/Pubmed Full Text
Geng, JJ, and Behrmann, M. (2002). Probabilità cuing della posizione di destinazione facilita la ricerca visiva implicitamente in partecipanti normali e pazienti con abbandono emispaziale. Psicol. sic. 13, 520–525.
Pubmed Abstract/Pubmed Full Text
Geng, JJ, and Behrmann, M. (2005). Probabilità spaziale come spunto di attenzione nella ricerca visiva. Percetto. Psicofisi. 67, 1252–1268.
Pubmed Abstract/Pubmed Full Text
Geng, JJ, e Mangun, GR (2011). L’attivazione della giunzione temporoparietale destra da un segnale contestuale saliente facilita la discriminazione del bersaglio. Neuroimage 54, 594-601.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Gibson, B. S., and Kelsey, E. M. (1998). La cattura attenzionale guidata dallo stimolo è subordinata al set di attenzione per le funzionalità visive displaywide. J. Scad. Psicol. Ronzio. Percetto. Eseguire. 24, 699–706.
Pubmed Abstract/Pubmed Full Text
Giesbrecht, B., Woldorff, M. G., Song, A. W., e Mangun, G. R. (2003). Meccanismi neurali di controllo dall’alto verso il basso durante l’attenzione spaziale e caratteristica. Neuroimage 19, 496-512.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Gnadt, JW, and Andersen, R. A. (1988). Attività di pianificazione motoria correlata alla memoria nella corteccia parietale posteriore del macaco. Scad. Cervello Res. 70, 216-220.
Pubmed Abstract/Pubmed Full Text
Gottlieb, J., and Balan, P. (2010). Attenzione come decisione nello spazio informativo. Tendenze Cogn. Sic. 14, 240–248.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Greenberg, AS, Esterman, M., Wilson, D., Serences, JT, and Yantis, S. (2010). Controllo dell’attenzione spaziale e feature-based nella corteccia frontoparietale. J. Neurosci. 30, 14330–14339.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Hopfinger, JB, Buonocore, MH, e Mangun, GR (2000). I meccanismi neurali del controllo dell’attenzione dall’alto verso il basso. NAT. Neurosci. 3, 284–291.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Ipata, A. E. Nel 2006 è stato pubblicato il primo album in studio del gruppo. Le risposte delle labbra a uno stimolo popout sono ridotte se viene apertamente ignorato. NAT. Neurosci. 9, 1071–1076.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Johnston, W. A., and Dark, VJ (1986). Attenzione selettiva. Annu. Rev. Psicol. 37, 43–75.
Juola, J. F., Koshino, H., e Warner, C. B. (1995). Compromessi tra effetti attenzionali di segnali spaziali e sbalzi improvvisi. Percetto. Psicofisi. 57, 333–342.
Pubmed Abstract/Pubmed Full Text
Kastner, S., Pinsk, M. A., De Weerd, P., Desimone, R., e Ungerleider, L. G. (1999). Aumento dell’attività nella corteccia visiva umana durante l’attenzione diretta in assenza di stimolazione visiva. Neurone 22, 751-761.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Koechlin, E., Ody, C. e Kouneiher, F. (2003). L’architettura del controllo cognitivo nella corteccia prefrontale umana. Scienza 302, 1181-1185.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Koshino, H., Warner, C. B., e Juola, J. F. (1992). Efficacia relativa di segnali centrali, periferici e improvvisi nell’attenzione visiva. Q. J. Scad. Psicol. A 45, 609-631.
Pubmed Abstract/Pubmed Full Text
Liu, T., Slotnick, S. D., Serences, J. T., and Yantis, S. (2003). Meccanismi corticali di controllo attenzionale basato sulle caratteristiche. Cereb. Cortex 13, 1334-1343.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Marois, R., Leung, H. C., and Gore, J. C. (2000). Un approccio stimolato all’elaborazione dell’identità e della posizione degli oggetti nel cervello umano. Neurone 25, 717-728.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
McFire, J., and Zangwill, O. L. (1960). Disabilità visuo-costruttive associate a lesioni dell’emisfero cerebrale sinistro. Cervello 82, 243-260.
Moore, C. M., e Egeth, H. (1998). In che modo l’attenzione basata sulle funzionalità influisce sull’elaborazione visiva? J. Scad. Psicol. Ronzio. Percetto. Eseguire. 24, 1296–1310.
Pubmed Abstract/Pubmed Full Text
Moran, J., e Desimone, R. (1985). L’attenzione selettiva porta l’elaborazione visiva nella corteccia extrastriata. Scienza 229, 782-784.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
O’Craven, K., Rosen, B. R., Kwong, K. K., Treisman, A., and Savoy, R. L. (1997). L’attenzione volontaria modula l’attività di fMRI in MT-MST umano. Neurone 18, 591-598.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Piercy, M. (1964). Gli effetti delle lesioni cerebrali sulla funzione intellettuale: una revisione delle attuali tendenze della ricerca. Fr. J. Psichiatria 110, 310-352.
Pubmed Abstract/Pubmed Full Text
Posner, M. I. (1980). Orientamento dell’attenzione. Q. J. Scad. Psicol. 32, 3–25.
Pubmed Abstract/Pubmed Full Text
Posner, MI, Snyder, CR e Davidson, BJ (1980). Attenzione e il rilevamento di segnali. J. Scad. Psicol. 109, 160–174.
Pubmed Abstract/Pubmed Full Text
Posner, M. I., Walker, J. A., Friedrich, F. J., e Rafal, R. D. (1984). Effetti della lesione parietale sull’orientamento nascosto dell’attenzione. J. Neurosci. 4, 1863–1874.
Pubmed Abstract/Pubmed Full Text
Serences, JT, Shomstein, S., Leber, AB, Golay, X., Egeth, H. E., e Yantis, S. (2005). Coordinamento del controllo attentivo volontario e stimolato nella corteccia umana. Psicol. Sic. 16, 114–122.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Shaw, ML (1978). Un modello di allocazione della capacità per il tempo di reazione. J. Scad. Psicol. Ronzio. Percetto. Eseguire. 4, 586–598.
Shomstein, S., e Behrmann, M. (2006). Sistemi corticali che mediano l’attenzione visiva sia agli oggetti che alle posizioni spaziali. Proc. Natl. Acad. Sic. U. S. A. 103, 11387-11392.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Shomstein, S., Lee, J. e Behrmann, M. (2010). Guida attentiva dall’alto verso il basso e dal basso verso l’alto: indagare il ruolo delle cortecce parietali dorsali e ventrali. Scad. Brain Res. 206, 197-208.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Shomstein, S., and Yantis, S. (2004a). Priorità configurale e contestuale nell’attenzione basata sugli oggetti. Psychon. Toro. Apoc 11, 247-253.
Pubmed Abstract/Pubmed Full Text
Shomstein, S., and Yantis, S. (2004b). Il controllo dell’attenzione si sposta tra visione e audizione nella corteccia umana. J. Neurosci. 24, 10702–10706.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Shomstein, S., and Yantis, S. (2006). La corteccia parietale media il controllo volontario dell’attenzione uditiva spaziale e non spaziale. J. Neurosci. 26, 435–439.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Silver, M. A., Ress, D., e Heeger, DJ (2005). Mappe topografiche dell’attenzione spaziale visiva nella corteccia parietale umana. J. Neurofisiolo. 94, 1358–1371.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Tamber-Rosenau, BJ, Esterman, M., Chiu, Y.-C. e Yantis, S. (2011). Meccanismi corticali di controllo cognitivo per spostare l’attenzione nella visione e nella memoria di lavoro. J. Cogn. Neurosci. 23, 2905–2919.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Theeuwes, J. (1991). Controllo esogeno ed endogeno dell’attenzione: l’effetto di onsets visivi e offset. Percetto. Psicofisi. 49, 83–90.
Pubmed Abstract/Pubmed Full Text
Vallar, G., and Perani, D. (1986). L’anatomia dell’abbandono unilaterale dopo lesioni da ictus dell’emisfero destro. Uno studio di correlazione clinica/CT-scan nell’uomo. Neuropsicologia 24, 609-622.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Wojciulik, E., and Kanwisher, N. (1999). La generalità del coinvolgimento parietale nell’attenzione visiva. Neurone 23, 747-764.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Yantis, S., and Jonides, J. (1984). Sbalzi visivi bruschi e attenzione selettiva: prove dalla ricerca visiva. J. Scad. Psicol. Ronzio. Percetto. Eseguire. 10, 601–621.
Pubmed Abstract/Pubmed Full Text
Yantis, S., and Jonides, J. (1990). Sbalzi visivi bruschi e attenzione selettiva: allocazione volontaria rispetto a quella automatica. J. Scad. Psicol. Ronzio. Percetto. Eseguire. 16, 121–134.
Pubmed Abstract/Pubmed Full Text
Yantis, S., Schwarzbach, J., Serences, J. T. Nel 2002 è stato pubblicato il primo album in studio del gruppo. Attività neurale transitoria nella corteccia parietale umana durante i cambiamenti di attenzione spaziale. NAT. Neurosci. 5, 995–1002.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Yantis, S., and Serences, JT (2003). Meccanismi corticali di controllo attenzionale basato sullo spazio e sugli oggetti. Curr. Opin. Neurobiolo. 13, 187–193.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text