広告:
この記事では、バッタについて説明します:-1。 バッタの習慣、生息地および外部の特徴2。 バッタの内部解剖3. 消化器系4. 第5話では登場していない。 呼吸器系6. 排泄システム7. 神経系8. Sense Organs9. 生殖システム10。 経済的重要性11. コントロール。
コンテンツ:
- バッタの習慣、生息地と外部の特徴
- バッタの内部解剖学
- バッタの消化器系
- バッタの循環系
- バッタの呼吸器系
- バッタの排泄系
- バッタの神経系
- バッタの感覚器官
- バッタの感覚器官
- バッタの感覚器官
- バッタの感覚器官
- バッタの感覚器官
- バッタの感覚器官
- バッタの感覚器官
- バッタの感覚器官
- バッタの感覚器官
- バッタの感覚器官
- バッタの感覚器官
- バッタの感覚器官
- バッタの感覚器官>バッタの生殖システム
- バッタの経済的重要性
- バッタの制御
バッタの習慣、生息地と外部の特徴:
バッタは世界的に分布しており、開いた草原と豊富な緑豊かな植生がある場所で発見されています。 彼らは葉の多い植生を食べる。 彼らは本質的に孤立した居住種であり、しばしば個体として豊富であるが、時折移動する可能性がある。
広告:
イナゴは群生し、渡り鳥の形です。 時には、イナゴが大量に増加し、群れで長距離を移動し、攻撃し、作物や植生に計り知れない損傷を引き起こすことがあります。

バッタの外部機能:
(i)形状とサイズ:
広告:
バッタの体は狭く、細長い、円筒形で、左右対称である。 それは長さが8cmまで測定する比較的大きい昆虫です。
(ii)色付け:
通常の体の色は黄色く、茶色がかった色で、マーキングや色の斑点が異なります。
(ii)色付け:
通常の体の色は黄色く、茶色がかっ キチンの顔料は環境に一致させるボディに保護着色を提供します。

(iii)外骨格:
体は、内の繊細なシステムを保護する外骨格で覆われています。
(iii)外骨格:
体 この外骨格は、キチンからなり、セグメントの線形の列に分割されているキューティクルです。 外骨格は、体のセグメントと付属物の動きを可能にする柔らかいキューティクルによって分離された硬いプレートまたは硬化物に形成される。
より柔らかい領域は縫合糸として知られています。 各部分はscleritesとして知られている別の部分から成っています。 通常典型的な区分のscleritesのいくつかは区別することができないし、従って縫合線は時代遅れまたは不明瞭であると言われます。 体壁は、それを分泌する皮下組織の細胞層であり、この基底膜の下にあるクチクラで構成されています。
広告:
(iv)バッタの体の分割:
広告:
バッタでは、体は三つの典型的な領域、すなわち。、頭部、胸郭および腹部。
(A)ヘッド:
バッタの頭は多かれ少なかれ腹側ですが、摂食中には認知度が低いように見えます。 それはキチン質のカプセルに封入されており、子宮頸部硬化物を有する小さな首によって体に取り付けられている。 六つのセグメントは、頭を形成するために一緒に融合されています。 頭部は背側の部分、頂点から成っています;前部の領域、frons;および側面、またはgenae。
フロンの下にはプレート、クリペウスがあります。 頭の両側に複眼があります。 三つの単純な目またはocelliは複眼の間の領域に位置しています。 頭部には一対の細い触角も見られる。 頭の腹側には口の部分があります。 p>

頭の付属物:
頭は一対の触角、一対の複眼、三つの単純な目またはocelliと口の部分を負います。
(b)触角:
触角は糸状です。 各アンテナは、花茎と呼ばれる小さな部分、未分化の小花茎、および約二十から五セグメントからなる十分に長い鞭毛で構成されています。 触角の表面には感覚的な剛毛、おそらく本質的に嗅覚が存在する。
広告:
(c)複眼:
二つの複眼は、頭の最初のセグメントに背側に配置されています。 茎がないので、これらは固着しています。 各複眼はキューティクルの透明な部分で覆われており、角膜は多数の六角形のファセットに分割されています。 各ファセットはommatidiumとして知られている単位の外の端である。 このような構造はモザイクビジョンを与える。
(d)オチェリ:
三つの単純な目またはocelliは、複眼の間に配置されています。 眼窩は、網膜とキューティクルの修正である厚い透明なレンズの視覚細胞のグループで構成されています。
(e)口の部分:
バッタの口の部分は咀嚼型または下顎型である。
広告:
口の部分には、唇、下顎、上顎、唇および下咽頭が含まれます。 口唇の腹側の縁には、唇または上唇が付いています。 この下には膜状の舌のような器官、下咽頭があります。 どちらかの側面で粉砕のための歯付き表面の付属品が付いている単一、堅い顎または下顎はある。 下顎骨の下には一対の上顎骨があります。
各上顎は、基底カルド、中央の茎、長い湾曲したラシニア、長い丸みを帯びたガレアおよびpalpiferから生じる上顎palpで構成されています。 陰唇または下唇は、基底の下腹部、中央のメンタム、二つの可動フラップ、ligulaeと両側の唇palpを備えています。
(f)胸郭:
胸郭は、柔軟な関節によって頭と腹部から分離されています。
それは三つのセグメントで構成されています:
前胸郭、中胸郭および後胸郭。 これらのセグメントのそれぞれには一対の脚があり、中胸郭と中胸郭はそれぞれ一対の翼を持っています。 中胸郭と中胸郭の両側には、呼吸器系への開口部である螺旋があります。
広告:
典型的なセグメントには十一の硬化物が含まれています。 背側帯状疱疹(それぞれ前胸、中胸、中胸、中胸で前胸、中胸、中胸と呼ばれる)は、四つの強膜からなり、前前胸、後胸膜、後胸膜に続いている。 側胸膜は三つの強膜,エピテルナム,エピメロン,パラプテロンからなる。 単一の腹側硬化物は胸骨である。
前胸郭の前鼻は大きく、両側に伸びています。 胸骨には背骨があります。 中胸筋では中胸筋は小さいが胸膜の硬膜は明瞭である。 中胸郭の硬化物は中胸郭に似ています。 p>

(g)脚:
各胸部セグメントは、接合された脚のペアを負います。
各脚は、次のように五つのセグメントの線形シリーズで構成されていますコクサは、体と関節し、大腿骨、脛骨および足根と融合した小さな転子が来ます。 各脚の足根は三つの目に見えるセグメントで構成され、脛骨に隣接するものは腹側表面に三つのパッドを有し、末端セグメントは、肉質の葉、pulvillusである間の爪のペアを負います。 p>

(h)翼:
バッタでは、中胸と中胸のそれぞれが一対の翼を持っています。 前翅は狭く、主に羊皮紙のようなものです。
これらは均一にまたは基盤の方の深まる陰と着色されるか、または斑点を付けるかもしれない。 このペアは、休息の位置では腹部と後肢の翼を覆うため、tegminaとも呼ばれます。 後羽は広く膜状であり、交互の方向に多数の縦方向の折り目を有する扇状の方法で折り畳まれて保持される。
各翼は、体の覆いの嚢のような突起として発達し、気管、神経および血液洞を囲む薄い二重膜に平らになる。 キューティクルは副鼻腔に沿って厚くなり、神経や静脈を強化します。 これらの静脈は異なる種の間でそのパターンが異なるが、それらは分類のために役立つ特定の種の個体において一定である。
(i)腹部:
腹部は細長く、後端に向かって先細りであり、末端セグメントは交尾または産卵のために特化されている。
(i)腹部:
腹部は後端に向かって先細りである。 それは11のセグメントで構成されています。 各セグメントは、典型的には背側tergumと腹側胸骨を有し、胸膜は存在しない。 腹部の第一セグメントの胸骨は胸郭に融合され、そのtergumは聴覚嚢を覆う楕円形の鼓膜の両側にある。
終末セグメントは、交尾と産卵のために男女の両方で修正されています。 男性では腹部の端は丸みを帯びていますが、女性では尖っています。 男女ともに、9と10のセグメントのtergaは部分的に融合しています。 男性では、11セグメントのtergumは、肛門の上に肛門上の肛門板を形成する。
10セグメントと9セグメントの胸骨の後ろにそれぞれの側にcercusプロジェクトと呼ばれる小さなプロセスは長く、二つの短い突起で背側に終わ 副生殖器プレートは、男性生殖器装置を覆う。
女性では、9セグメントの胸骨は細長く、腹部は2対のローブまたは弁で終わり、より大きな弁の間に隠されたより小さな対がある。 Ovipositorはこれらの3組の弁から成っています。 8対の螺旋が存在し、1つの螺旋は2から9のセグメントのいずれかの下側に位置しています。
バッタの内部解剖学:
バッタの内部腔は、血液を含み、真の腔腔ではない、すなわちhaemocoelである。 器官のシステムはhaemocoelの内にあります。 p>

筋肉系バッタ:
筋肉は横紋型で、非常に柔らかく繊細ですが強いです。 筋肉の数は非常に多いです。 それらは腹部に分節的に配置されるが、頭部および胸部には配置されない。 最も顕著な筋肉は、下顎骨、翼、中胸骨脚および産卵管を動かす筋肉である。
バッタの消化器系:
バッタの消化管:
バッタの消化管は、三つの主要な領域、すなわちで構成されています。、前腸、中腸および後腸。
前腸またはstomodaeumは、口の部分に囲まれた口から始まり、非常に短い筋肉の咽頭に開きます。 咽頭は短く、狭く、細く、管状の食道につながり、これは拡張された円錐嚢のようで薄壁の構造に拡大し、作物は胸郭の後端まで伸びる。
作物は突然膨張して、proventriculusまたは砂嚢と呼ばれる厚い硬いわずかに円錐形の構造を形成する。 Proventriculusは歯を支えるいくつかの堅いキチン質の版を作動させる大きく強力な円筋肉の存在のために厚い壁である。 この複雑な構造は重要な咀嚼装置であり、固形食品を粉砕する。
proventriculusの後端にある大きな括約筋は、中腸への通過を制御するために心臓弁を形成する。 一対の小さな分岐唾液腺が食道および作物の腹側に付着していることが見出される。 唾液腺の管は、陰唇の口腔内に開きます。 前腸はキチン質の内膜と内部的に並んでいます。 中腸または腸間膜は、心室または胃である。
これは、心臓括約筋からMalpighian尿細管の起源のポイントに四から五腹部セグメント内に位置する非常に顕著なほぼ直線管です。 その透明な膜状の壁は、前腸のようにキューティクルで裏打ちされていません。
六つの二重指状の肝caecaまたは胃caecaのシリーズは、二つのグループでその前端から発生します。 これらは、中腸の前端に独立して開く。
第六腹部セグメントに位置する幽門括約筋は、腸間膜の後端を示す。 多数の細い糸状の淡黄色のMalpighian細管がこの領域から起源を取り、haemocoelomic腔に独立して浮かんでいるのが見られることがあります。 p>

後腸または直腸は、拡大した前部、回腸、狭い中間部、結腸およびわずかに拡張されたが非常に薄い壁の顕著な直腸からなり、肛門によって外側に開口する。 後腸はキチン質の内膜によって内部的に裏打ちされています。
バッタの摂食と消化:
バッタは野菜の食べ物を食べる。 食べ物は前肢、labrumおよびlabiumによって保持され、唾液分泌(いくつかの酵素を含む)によって潤滑され、下顎骨および上顎骨によって噛まれる。 噛んだ食べ物は作物に保存されています。 それは砂嚢に少しずつ進み、そこでさらに粉砕され、緊張して胃の中に渡されます。
胃と肝臓caecaの壁にある腺は、消化をもたらすいくつかの酵素を分泌します。
胃と肝臓caecaの壁にある腺は、消化をもたらすいくつかの酵素 Midgutのわずかにアルカリか酸性分泌はマルターゼ、リパーゼ、ラクターゼ、プロテアーゼ、トリプシンおよびerepsinを含んでいます。 食品材料の吸収は中腸で起こる。
食品材料が直腸に到達するまでに、最大栄養材料が利用され、余分な水分が直腸に吸収される。
食品材料が直腸に到達するまでに、最大栄養材料 消化されていない物質または残留物は、肛門を通って排出される細い糞便ペレットに変換される。
バッタの循環系:
毛細血管や静脈がないため、循環系は開いたもの(ラクナー)です。 それは他の多くの節足動物と比較してはるかに減少しています。 腹部に中央背側に横たわっている単一の、細身の、管状および拍動性の心臓がある。 それはtergitesの凹面の内部の表面を渡って伸びる敏感な横断ダイヤフラムによって形作られる浅い心膜キャビティで中断される。
心臓には、オスティアという名前の側開口部がいくつかあり、一方向のみの血液の流れを可能にする弁が設けられています。 いくつかの筋肉繊維は収縮および弛緩によって心膜のキャビティを拡大し、減らすために、alary筋肉横隔膜に発散的にファンワイズ広がっています。
一方の筋肉の繊維は、心臓の下の反対側の対応する筋肉の繊維と一致します。
Alary筋肉の付着点の間には、両側に空間があり、それを通って血液腔からの血液が心膜洞に入る。
横隔膜上の収縮の波が閉じ、間隔が開き、オスティアの弁を押して血液を心臓に送り込む。 この構造の脈動では、血液は長い背側大動脈を通って頭部領域に前方に流れ、血液循環腔に戻る。
解剖学的には、心臓および大動脈は、心臓の室として知られる管の部分的な拡張の存在によって区別することができる。 バッタには一般的にそのような七つの部屋があります。 オスティアは、したがって、それらの側壁に三日月状の開口部である。 大動脈は背側血管の胸部であり、胸郭を通過した後、頭部に入る。

背側横隔膜のほかに、胸郭前から体の端まで連続したシートを形成し、その下に神経周囲洞を囲む腹側横隔膜がある。 閉鎖された静脈か毛管がほしいと思っているが、血は付属物および翼の静脈にすっかり循環します。
血漿には、異物を除去する食細胞として作用する無色の血液細胞が含まれています。
血漿には、異物を除去する食細胞として作用する無色の血 血液は、主に食品や廃棄物を輸送するのに役立ちます。 脂肪体は、様々な器官および神経索を完全に包み込む黄色の細胞の緩く凝集した塊からなり、多かれ少なかれ鞘として作用する。 それらは不利な条件の下で使用のための食糧を貯える。
バッタ(イナゴ)の呼吸器系:
呼吸器系は、体のあらゆる部分と通信する気管である外胚葉管のネットワークで構成されています。 気管は、細胞の単層からなり、キューティクルで裏打ちされています。 最も大きい気管の管はキチン、それらが倒れることを防ぐtaenidiaの螺線形の糸を所有しています。
体の両側の螺旋は、枝によって長手方向の幹につながる。 最高級の気管である気管孔は、酸素を供給し、二酸化炭素を運ぶために身体組織に直接接続されています。 筋肉や他の器官の気管の小さな盲目の終わりは、液体で満たされています。 筋肉の活動中、気管周囲の体液中の物質の濃度が増加する。
これは、気管から周囲の領域への水の拡散を引き起こし、空気が気管の盲目の先端に遠くに移動するにつれて、酸素が使用されている部位に近 活動が停止した後、浸透圧を変化させた代謝産物が処分され、水は気管に戻る。
腹部には、腹部の交互の収縮と拡張によって気管系の内外に空気を送り込むいくつかの薄肉気嚢もあります。 バッタでは、螺旋の作用は非常に同期しており、螺旋の最初の四つのペアはインスピレーションで開き、満了時に閉じられ、他の六つのペアはインスピレーションで閉じ、満了時に開いている。 p>

バッタの排泄システム:
排泄器官は、haemocoelの周りに巻かれ、後腸の前端に開いているMalpighian尿細管です。 Malpighian尿細管は、線条状の内側の境界を有する細胞の単層の壁を有する。 彼らの自由端は完全に閉じられています。
血液からの代謝廃棄物は、Malpighian尿細管の細胞によって抽出され、尿細管の内腔に渡され、最終的に肛門を通って排出されるために腸に排出される。 Malpighian尿細管はhaemocoelにあるので、尿酸、尿素、尿酸塩、炭酸カルシウムおよびシュウ酸塩および塩を取除く。 p>

このページのトップへ:
脳または食道上神経節は、食道の上の頭部に背側にある。 それは目、触角およびlabrumに神経を与える3組の融合された神経節(protocerebrum、deutocerebrumおよびtritocerebrum)から成っています。
脳は、再び神経節の三対の融合によって形成されたサブ食道神経節に二つのスタウトcircumoesophageal連結体によって結合されています,すなわち.、下顎、上顎および唇。 それは頭部の中央の口の部分の上に位置し、後部側にわずかに傾斜しています。 この8対の神経から、下顎、上顎、陰唇、下咽頭、頸部、頭部および唾液領域に放出される。
食道下神経節からは、対になった神経節と縦連結体からなる腹側神経索が後方に延びている。 各胸部の区分は足、翼および内臓に神経を供給する組の神経節を含んでいます。 様々な後部器官に神経を送る腹部神経節の唯一の五対があります。
内臓または交感神経系もあり、食道部分または胃胃神経系で構成され、神経節および神経が脳に接続して腸の前部に供給され、腹側交感神経系が後腸および生殖器系に神経を供給する。 末梢神経の細かいパターンは、体壁の表皮の下にあります。 p>

バッタの感覚器官:
バッタの感覚器官は、空気やそれが住んでいる他の環境からの刺激を受け、動きや他の応答による外部の変化を調整するために適応されてい
これは、運動組織を制御する神経管のメカニズムを介して中枢神経系に伝達される外部刺激を受信するためにsensillaと呼ばれる特に設計された構造を形成する体壁の特殊な細胞の開発によって達成されます。 感覚器官は体表面に広く分布しており、付属器は消化管の前部および後部にも発生しています。
グラスホッパー内では、以下の感覚器官が満たされています。
1。 触覚器官:
彼らはsetae、棘、毛、コーン、剛毛などの形をしています。 体の様々な部分、特に触角、口の部分、脚、翼、生殖器などに散在しています。 触覚器官は接触に敏感である。
2. 嗅覚器官:
嗅覚器官は匂いに敏感です。 触角には嗅覚の主要な器官が供給されています。
3. 味覚器官:
味覚の器官は、嗅覚器官に似た形で、口の部分に特にpalps、咽頭、触角、およびtarsiに発生します。
4. 視覚器官:
バッタは大きな複眼と三つのocelliのペアを持っています。 複眼は視覚と光の知覚のためのocelliに関係しています。 眼窩は、光受容体細胞または網膜のグループで構成され、それぞれが脳につながる神経線維で終わる。 各光受容体の外側端はラブドームを形成する。
光受容体細胞の群を覆うキューティクルは、厚い両凸、透明なレンズを形成する。 Ocelliの実際の機能は明確には分かっていません。 複眼は、構造的にも機能的にもゴキブリ、エビ、ザリガニのものに似ています。
5. 聴覚器官:
バッタはストライドリング装置で特定の音を作り出すので聞くことができると考えられています。 聴覚器官のペアは、最初の腹部セグメントのtergiteの側面に位置しています。 各聴覚器官は、ほぼ円形のキチン質のリング内に伸びた鼓膜または鼓膜からなる。 それは空気中の音の振動によって動きに設定されます。
それは空気中の音の振動によって動きに設定されます。 これはそれから感覚神経繊維に接続される膜の下の細いポイントに影響を与えます。 p>

バッタの生殖器系:
男女は別々であり、男性と女性のバッタの区別は腹部の後端によって決定することがで オスでは丸く、メスでは産卵管のために尖っている。
男性の生殖器官:
男性の生殖器官(図。 74.14A)は、二つの精巣、二つのvasa deferentia、二つの精嚢、単一の射精管、単一の陰茎および副腺のペアで構成されています。 両方の精巣は、腸の上の脂肪体の塊に埋め込まれている。 各精巣は、精子が発達する一連の細い細管または卵胞で構成されている。
精管と呼ばれる複雑な管が各精巣から導かれます。 各精管は、精嚢と呼ばれる嚢のような構造に後方に拡張されています。 それはposteriorly狭くし、自身の側面の頑丈なthick-walled顕著な付属腺に会う。
精嚢および副腺の大きさおよび形状は、異なる種で異なる。 どちらかの側から来る2つの精嚢は共通の中央の射精管を形作る一緒に会います。 このダクトは、大きな腹側の男性の交尾器官、陰茎またはaedeagusの終わりに開きます。 副腺は、交尾中に精子を女性に移すのに役立つ液体を分泌するように見える。
女性の生殖器官:
女性の生殖器官(図。 74.14B)一対の卵巣、卵管、付属腺、中央膣、および精嚢または精嚢を含む。 各卵巣は、いくつかの卵管または卵巣細管で構成されており、その中で多数の卵子が産生される。 各ovarioleは先細りの管であり、その壁の厚さは後方に増加する。
卵子は内腔に流され、サイズが成長するにつれて後方に下降するので、この管は発達の様々な段階で卵子の寸法が異なるため、ビーズの外観を与える。 Ovariolesの末端は絡み合っています。 後卵管は、共通のダクト、卵管を形成するために一緒に会います。
両側からの卵管は、わずかに厚く、筋肉である中央の短い膣を形成するために一緒に会います。 それはposteriorly動き、ovipositorの版の間で腹側に開く。 一対の顕著な付属腺は、膣と独立して会合する。
小さな嚢、精嚢または精嚢は、小さな狭い管によって膣に結合する。 交尾の間に精液は精液の容器で受け取られ、貯えられます。 彼らは膣領域を通過するときに卵を受精させる。

交尾:
交尾は夏の終わりに発生します。
交尾は夏の終わりに発生します。
交尾は夏の終わりに発生します。
交尾は夏の終わりに発生します。
交尾は夏の終わりに 交尾では、男性のバッタは女性の背中にしがみつき、陰茎を膣に挿入し、精子を移す。 精子は、卵が産生されるまで精液容器に貯蔵される。 交尾は、女性が卵を産み始める前に数回行われることがあります。

産卵
受精:
成熟した卵、3-5mmの長さは、卵管を通過します。 各卵は敏感な内部のvitellineの膜および精液が置くことの間に入り、卵を受精させる微細な気孔かmicropyleを含んでいる茶色がかった適用範囲が広い貝か絨毛膜 精子核は成熟した卵の核と結合し、胚が発生する卵の周囲に胚葉が形成される。

産卵:
産卵は交尾後に短い間隔を開始し、秋に続きます。 女性は卵を堆積させ、卵ポッドとしてそれらを一緒に固定粘着性の分泌物に囲まれている地面に短いトンネルや穴を形成するために彼女のovipositorを使 卵は通常、二十の多くに敷設されており、単一の女性は十ロットまで産むことができます。 成虫は交尾と産卵の数日後に死ぬ。
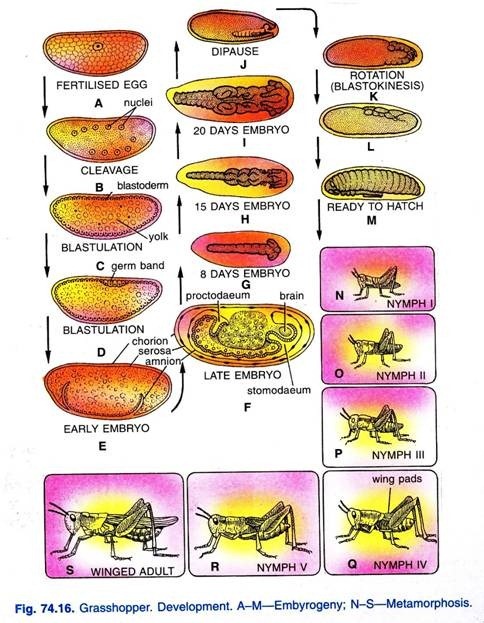
開発:
胚発生(図。 74.16)は、胚がよく形成されるまで約三週間続きます。 その後、発達が阻止され、胚は冬の寒さと食物不足の不利な条件を乗り越えるために、休息期間、または休眠期に入ります。 成長は、温度が暖かい春に再び始まります。
卵から孵化する若いバッタはニンフと呼ばれています。 それはその親に似ていますが、体の残りの部分と比較して大きな頭を持っており、翼と生殖器官を欠いています。 それは植生に餌を与え、急速に成長する。 若いバッタが成長し、定期的に流され、その柔軟性のないキチン質の外骨格のために大きすぎるようになります。
キチン質の外骨格の脱落は、脱皮と呼ばれる複雑なプロセスです。 翼は翼パッドから徐々に発達し、5つの脱皮の後、若いバッタは成体の形に達する。 このタイプの開発は、単純または漸進的な変態と呼ばれています。 p>
バッタの経済的重要性:
(i)作物害虫として:
ニンフと大人の両方が多くの種類の植生、特に多肉植物の種類を食べる。 彼らはしばしば新しい餌場に移動し、農場や庭の植え付けを損傷したり台無しにする可能性があります。 摂食は、かなり晴れた日の午前中に最も活発です。 食糧が不足しているとき、これらの昆虫は綿や毛織物、木や障害バッタを食べるでしょう。
バッタはまた、草を食べ、したがって、範囲や牧草地に大きな被害を与えます。 また、長い大群に移行バッタである真のイナゴは、異なる種類のものであり、私たちの作物のフィールドや他の植生に大きなダメージを与えます。 東半球で発見された回遊性イナゴ、イナゴmigratoriaは、聖書の時代から飢饉を引き起こしています。
Melanoplus maxicans、北アメリカのロッキーマウンテンイナゴはまた、大きな損失を引き起こすM.maxicans spredusと呼ばれる移動期を生じさせる。
バッタCamnula pellucidaは深刻な害虫です。 好条件と敵の不足の間に、それは5月または6月に発達し、孵化します。 それは渡り鳥の形であり、長い距離を飛ぶことができます。 このバッタの群れは緑の植生を破壊し、エジプト人によってバッタの疫病と呼ばれていました。 (ii)食べ物として:
バッタはまた、人間や他の動物にいくつかの使用されています。
(ii)食べ物として:
バッタは、人間や他の動物にもいくつかの使用されています。 彼らは生きているか死んでいるかのどちらかの良い魚の餌として使用されます。 彼らは時には人間の食べ物にも使用されます。 日本、メキシコ、フィリピンなどの国では、まだ食品として使用されています。 彼らは一般的に北米インディアンと世界の他の部分で原始的な部族によって食べられています。
ギリシャ人は迫撃砲でイナゴを粉砕し、それらの小麦粉を作り、小麦粉を食糧として使用しました。 バッタの卵、ニンフおよび成虫は、いくつかの捕食性昆虫、クモ、カエル、爬虫類、鳥類および哺乳動物のための食物を提供する。 インドでも一部の人々は、ローストや揚げ食品としてそれらを食べます。
バッタの制御:
バッタは、自然だけでなく、人工的または化学的手段によって制御されています。
バッタの制御:
バッタは、人工的または化学的手段によって制御されています。 バッタの卵は、いくつかのカブトムシ、蜂のハエ、モル、スカンク、マウス、強盗のハエと掘りスズメバチとニンフと大人の両方の大きな捕食昆虫とカエル、爬虫類、鳥や哺乳類によって食べられています。 バッタの卵はまた、特定の昆虫に寄生されています。
肉ハエ(Sarcophaga)は成虫に生きているウジを産み、タキニドバエは飛行中にバッタに卵を預け、両方の幼虫は宿主に穴を開け、脂肪組織を消費する。 寄生されたバッタはlogyになり、再生するか、または死ぬために失敗します。
寄生昆虫は、このように、バッタ制御の要因を構成しています。 真菌と細菌の両方の病気はまた、時にはバッタの数を破壊します。 土が耕すことによって太陽にさらされると、バッタの卵は冬の間地面で殺されます。 昔、バッタの制御は、ヒ素またはいくつかの他の胃の毒と混合した食品を与えることによって行われました。 しかし、今では殺虫剤が一般的に使用されています。
様々な殺虫剤は、スプレーまたは粉塵の形で使用され、接触または食べられたときに殺す毒餌である。 最近採用されているいくつかの殺虫剤は、アルドリン、ジエルドリン、クロラダン、ヘプタクロルおよびトキサファンである。 Methoxychlorはまた保護の果物と野菜および牧草地分野のために人か家畜に有害である残余を残さないので、殺虫剤として今使用されます。 p>