イントロダクション
私たちの感覚環境との相互作用を成功させるには、トップダウンとボトムアップの二つの注意選択メカニズムの複雑なバランスが必要です。 マンゴーサラダのためのいくつかの必要な成分を拾うことを目標に、あなたの地元のスーパーマーケットの農産物通路に向かって、あなたが積極的に農産物の選択肢の多数の中で必要なすべての成分を検索するような自主的な目標指向、またはトップダウン、注意システムの展開を行っています。 ただし、携帯電話の着信音を聞く必要があります、それは最も可能性の高いあなたの注意をキャプチャし、検索を中断します。 このような中断は、ボトムアップ、または刺激駆動型の方法で発生し、刺激の単なる顕著さ、リングがあなたの環境内の他の音とは異なるという事実は、選択の価値があると判断します。 説明されたシナリオは、行動のための目標指向と刺激主導の選択の重要性を強調し、一方では”トンネルビジョン”を防ぎ、他方に集中する完全な無力トップダウンとボトムアップの選択
: Behavior
数十年の行動研究は、注意の分布が観察者の意図によってだけでなく、物理的刺激の顕著さによって制御できることを実証することに専念し トップダウンおよびボトムアップの注意配分に関する行動的証拠の多くは、他の場所で広くレビューされている(Johnston and Dark、1986;Egeth and Yantis、1997)。 要約すると、トップダウンの注意制御の効果を実証する研究は、注意が空間的な場所、特徴、物体などにうまく割り当てることができることを示してい(Eriksen and H Offman,1 9 7 2;Posner,1 9 8 0;Posner e t a l. る(Shaw,1 9 7 8;Moore and Egeth,1 9 9 8;Geng and Behrmann,2 0 0 2,2 0 0 5;Shomstein and Yantis,2 0 0 4a;Drummond and Shomstein,2 0 1 0)。 ボトムアップの注意配分を支持する証拠は、参加者がトップダウン検索に従事し、彼らの注意がタスク無関係な刺激に転用され、注意が特徴のシングルトンによって捕捉されることを実証する様々な注意捕獲パラダイムに依存している(unique item;Yantis and Jonides,1990;Theeuwes,1991;Folk et al. ら、2 0 0 2)および突然の発生(YantisおよびJonides、1 9 8 4;Theeuwes、1 9 9 1;Koshino e t a l. ら、1 9 9 2;Juolaら、1 9 9 3;J., 1995).
ほとんどの初期の研究は、トップダウンとボトムアップの注意選択の証拠を実証することに集中していましたが、最近の研究では、二つの注意選択シ この一連の調査は、タスクに関連する情報(例えば、サラダの成分)を効果的に選択するためには、そうでなければ目の前のタスクから注意をそらすであろうタスクに関連しない情報を積極的に阻害しなければならないという観察によって支えられている。 このロジックの裏返しは、タスク関連の情報に焦点を当てているものが少ないほど、より多くのキャプチャが続くということです。 観察者の注意状態は、どのような種類の情報、そしてどの程度まで、最終的に注意を引くかを予測することが実験的に示されている(Folk et al. ら,1 9 9 2,2 0 0 2;Bacon and Egeth,1 9 9 4;Gibson and Kelsey,1 9 9 8)。 例えば、Folk e t a l. (2002)は、赤い文字を検索するとき、その刺激が赤色であるか、何らかの方法で標的テンプレートと一致する場合、周囲の無関係な刺激によって観察者がよ オブザーバーのトップダウン制御設定は赤の特徴を検索するように設定されているため、赤の刺激は注意を喚起し、トップダウン制御を妨げる可能性が したがって、キャプチャタスクでは、注意探索戦略は、ターゲットの刺激特性とディストラクタとの間の類似度レベルを変化させることによって、互いに区別することができる。 ターゲットがディスストラクターに似ているほど、観察者が捕獲を避けることがより困難になります。トップダウンとボトムアップの選択における頭頂葉の役割
: 神経イメージング
様々な神経イメージング技術は、トップダウンとボトムアップの向きに頭頂皮質の関与のための強力な証拠を提供し、証拠は他の場所で広範囲にレビューされた(Corbetta and Shulman,2002,2011;Behrmann et al., 2004). 特定の場所、特徴、または物体に対応するためにトップダウンの合図の後に最も一般的に活性化される領域は、頭頂皮質の背側部分に沿って位置す そのような領域には、下頭頂小葉(IPL)、上頭頂小葉(SPL)と呼ばれる背内側領域、ならびに前頚回に沿ったより内側領域が含まれる(Yantis e t a l. ら,2 0 0 2;Giesbrecht e t a l. ら、2 0 0 3;Liu e t a l.,2003;Yantis and Serences,2003;図1). いくつかのトップダウンタスクは、頭頂皮質の背側領域、すなわち空間を含む領域に首尾よく係合することが示されている(Kastner et al. ら、1 9 9 9;Corbetta e t a l. ら,2 0 0 0;Hopfinger e t a l. ら、2 0 0 0;ShomsteinおよびBehrmann、2 0 0 6;ChiuおよびYantis、2 0 0 9;Greenberg e t a l. ら、2 0 1 0)ならびに注目の非空間的シフト(Giesbrecht e t a l. ら、2 0 0 3;Yantis and Serences,2 0 0 3;Shomstein and Yantis,2 0 0 4b,2 0 0 6;Tamber−Rosenau e t a l., 2011).
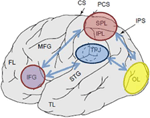
図1. 人間の脳の側面に投影された関連する解剖学的ランドマークの概略的な描写。 上頭頂小葉(SPL)および下頭頂小葉(IPL)は、頭頂皮質の背側部分内の領域であり、トップダウンの注意指向性を有する。 頭頂-頭頂接合部(tpj)は、ボトムアップの注意方向を支配する腹側頭頂皮質内の領域である。 SPLとTPJの両方が、感覚領域にわたって観察されたその後の注意変調の原因となる制御信号を引き出すと考えられており、この場合、後頭葉(OL)の視覚的に誘発された活動を変調する(濃い青色の矢印で標識されている)。 さらに、下前頭回(IFG)および下前頭接合部(IFJ)に沿った領域は、刺激駆動およびトップダウン注意制御(水色の双方向矢印でマーク)の収束領域として役立つと考
トップダウンの注意割り当てを行うことを目的とした典型的なタスクでは、個人は周囲に配置された二つの急速な連続視覚提示(RSVP)ストリームを示 キューの同一性は、被験者が現在の流れに注意を維持しなければならないか、または他の流れに注意をシフトしなければならないかを示す(Yantis et al. ら,2 0 0 2;YantisおよびSerences,2 0 0 3)。 このようなパラダイムでは二つの主要な知見が観察されている。 最初のものは、現時点で出席した場所を表す感覚領域内の活性化の増加(例えば、右のRSVPストリームが出席したときの左の一次視覚領域内の活性の増加) この知見は、参加者が特定の場所に出席しており、注意が感覚応答の強さを調節するという確固たる証拠を提供する(図1を参照;Moran and Desimone,1985;O’Craven et al., 1997). 第二の発見は、頭頂葉の背側領域がトップダウン注意のシフトによって選択的に活性化されるという観察に関係している。 活動のSPL/IPLタイムコースは本質的に一時的であることが観察され、頭頂皮質のこの領域が、注意深い状態をトップダウン的にシフトさせるための簡単な注意制御信号の源であることを示唆している(Yantis et al., 2002).
いくつかのfMRI研究では、刺激の顕著性および/または関連性によって媒介されるボトムアップの注意捕獲が、側頭頂接合部によって維持されるこ 例えば、被験者が同時に提示された視覚刺激または聴覚刺激のいずれかの変化に出席し、監視する場合、頭頂葉のTPJ領域の活性化が増強される。 関連する刺激に対する見かけの感受性に加えて、TPJは、生物が中立的な行動文脈に従事しているとき、またはタスクに従事しているときに、潜在的に新し ら,2 0 0 0;Downar e t a l. ら,2 0 0 2;Serences e t a l. ら、2 0 0 5;Corbetta e t a l. ら、2 0 0 8;Asplund e t a l. とができることを示唆している。 この活性化は、TPJの多感覚的性質を反映して、入力が送達されるモダリティ(聴覚、触覚、および視覚)とは無関係に起こる(ただし、Downar e t a l., 2001).
ボトムアップ注意捕獲の神経機構を調べる典型的なタスクでは、参加者はディスプレイの中央にアイテムのRSVPストリームを提示され、事前に定義されたターゲットを識別するように求められます(例えば、白い非ターゲットのRSVPストリーム内に提示された赤い文字を識別する)。 いくつかの割合の試験には、標的の発症前の様々な時間間隔で提示されたタスク無関係な突出した散漫因子が含まれていますが、他の試験には突出した散漫因子のみが含まれています(すなわち、標的なし)。 「Target−distractor」試験は、捕捉の程度をアッセイするために使用され、タスク無関係のdistractorが実際に顕著であり、それによって標的精度の低下をもたらすことを示す。 “Distractor-in-isolation”試験は、標的関連プロセスからの汚染なしに顕著なdistractorに誘発される活性の検査を可能にするため、さらなる分析に使用されます。 いくつかの重要な知見は、このようなパラダイムから出てくる。 まず、デストラクタがターゲット位置から空間的に分離されている場合、キャプチャデストラクタは、感覚皮質の対応する領域(例えば、網膜光学的に組織化された視覚野;図1を参照)における皮質活動の増加を伴う。 そのような結果は、捕捉の間、空間的注意が、実際には、散漫器によって占有される空間的位置に捕捉されるという強力な証拠を提供する(Serences et al., 2005). 第二に、主にTPJ内の頭頂皮質の腹側領域は、ボトムアップ、不随意、注意のシフトによって選択的に活性化される。 トップダウン指向のためのSPL内の活動と同じように、TPJ上で観察された活動のタイムコースは、この領域がボトムアップの方法で注意をシフトするた
このレビューは、後頭頂皮質の認知機能に対処することに焦点を当てていますが、他の領域、特に前頭皮質内の領域もトップダウンおよびボトムア そのような領域には、腹側前頭皮質(VFC)、前頭眼野(FEF)、下前頭接合部(IFJ)、および下前頭回が含まれる(IFG;CorbettaおよびShulman、2 0 0 2、2 0 1 1;Serences e t a l. ら、2 0 0 5;Asplund e t a l. ら,2 0 1 0;DiquattroおよびGeng,2 0 1 1)。トップダウンとボトムアップの選択における頭頂葉の役割
: 神経心理学
歴史的に研究者は、頭頂葉に関連する認知機能への洞察を得るために、半空間無視(左半空間への注意の空間的配分の障害)を有する患者の神経心理学的研究に批判的に依存していた。 古典的な神経心理学的文献では、頭頂皮質は、全体として、一般的に半空間無視のための原発病変部位と考えられていた。 この見解は、初期の研究者によって詳細に詳述されている(Critchley、1953;McFire and Zangwill、1960; Piercy,1964)は、頭頂病変とその後の無視との関連を明確に認識した。 この視点は、1980年代にポズナーら(1984年)が頭頂葉の損傷が、ターゲットが病変に対して対側であるときに”離脱”操作(ある場所から注意を引き込み、別の場所にシフトする)で欠損を生じることを示すために、秘密の視覚空間キューイングパラダイムを使用したときに主に保持されていた。 しかし、注意の神経基盤、特に頭頂皮質の”離脱”の役割を理解する上で、この大きな進歩にもかかわらず、彼らの知見は、単一の皮質部位(頭頂皮質)と単一の機能能(”離脱”)を前提としている。 脳(頭頂皮質)と行動(注意離脱)へのこのよりモノリシックなアプローチとは対照的に、最近の行動と神経イメージングの仕事(上記および他の場所でレビュー)は、皮質領域と関連する注意行動の両方が質的に異なるプロファイルに細分される可能性があることを示唆している。
皮質ネットワークのトップダウンとボトムアッププロセスへの分離を考えると、明らかな予測は、頭頂小葉の優れた部分への損傷(subsuming SPL)は、目標指向の注意指向の欠損をもたらすべきであるのに対し、頭頂小葉の劣った部分への損傷(subsuming TPJ)は、刺激駆動の注意捕獲に関連する欠損をもたらすであろうということである。 これらの脳と行動の対応が神経心理学の文献で探求されている限り、この予測は明らかに支持されていない。 例えば、片側性無視の臨床症状は、SPLのような優れた部分ではなく、tpjを含む頭頂葉の劣った部分への損傷と強く関連している(Friedrich e t a l. ら、1 9 9 8;Shomstein e t a l. ら,2 0 1 0;CorbettaおよびShulman,2 0 1 1)。 これは、tpjの役割は、ネグレクト患者が最も困難を持っているように見えるドメインではなく、注意の自発的な方向づけではなく、注意の捕獲にあることを示唆している神経画像化文献とやや対立している。 さらに問題を複雑にするために、SPLを排他的に含む病変は、無視の臨床的証拠を生じることはめったにないことが指摘されている(Vallar and Perani、1986)。 主にTPJおよびSTGを中心とする病変を有する患者を対象とした別の最近の研究では、SPLを保存していたが、Corbetta et al. (2005)は、空間無視とその回復が、腹側側頭頂および背側頭頂領域の両方における活動の回復と関連していることを示した(レビューについてはCorbetta and Shulman、2011を参照)。 興味深いとその結論で刺激的なが、この最後の研究は、患者が唯一のPosner秘密空間注意cuingタスク、トップダウンとボトムアップの両方の注意配向に係合
目標主導型の注意制御と顕著な注意捕獲を区別し、SPLとTPJへのマッピングを調べるために、最近の研究では、これらの注意の形態のいずれかをターゲッ, 2010). 患者におけるトップダウンの注意指向の完全性を調べるために、参加者が空間的に分離されたRSVPストリーム間で空間的な注意をシフトすることを必, 2002)). 同様に、患者のボトムアップの注意指向能力を調べるために、Folk et al. (2002)偶発捕捉パラダイムは、参加者が固定時に現れたターゲットを検出し、タスク無関係の色のシングルトンが周辺部でフラッシュされた。 タスクに無関係なディスクリプタが中央検出タスクに干渉する程度を、ボトムアップの注意捕獲の尺度として使用した(Bacon and Egeth,1994;Folk et al., 2002).
予測は次のとおりでした: 頭頂葉の上の部分への損害の患者(SPLに影響を与える)はトップダウンの注意の方向づけの仕事(捕獲の仕事の保存された性能と)で損なわれるべきであるが、頭頂葉の下の部分への損害の患者(TPJに影響を与える)は捕獲の仕事で損なわれるべきである(トップダウンの仕事の倹約された性能と)。 この形態の二重解離は、注意の独立した成分を証明するだけでなく、そのような注意成分が独立した神経機構によって媒介されることを示唆している。 Visuo空間無視を有する八人の患者は、研究のために募集され、刺激駆動または目標指向の注意指向のいずれかをタップし、二つのタスクを完了しました。 彼らの行動プロファイルに基づいて、患者はグループに分類され、それらの病変の重複が調査された(図2A)。 トップダウンの注意指数(図2B)によって定量化された目標指向の注意指向の困難を示した患者は、免れる下頭頂小葉(TPJ)と頭頂小葉(subsuming SPL)の上の部分を中心とした病変の重複を提示した。 病変が頭頂葉の下側部分を中心とした重複を有する患者(TPJを包含する)が、SPLは注意捕獲に対して免疫を維持しながら、目標指向の配向タスクで正常に行 この研究からの知見は、SPLとTPJは、必ずトップダウンとボトムアップ配向の目的のために募集されている解剖学的領域であり、SPLとTPJへの損傷は、それぞれトップダウンとボトムアップ配向の障害につながることを明らかに示唆している。
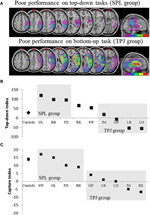
神経心理学的研究の結果は、トップダウンおよびボトムアップ方向へのSPLおよびTPJの相対的な寄与を調査することを目的とした。 (A)トップダウンの注意の方向づけの行動の赤字によってグループ化された患者の病変の重複(紫色の最小の重複;赤い最大の重複),SPLグループ(トップパネル); (B)トップダウンタスクの行動パフォーマンスは、左から右への空間的なトップダウンシフトとその逆の違いを定量化する”トップダウンインデックス”で要約されています。 対照群とTPJ病変群は空間シフトを実行する際に同様の効率を示し、SPL病変を有する患者は効率が低下したことを示した。 群対照および個々の患者データ(患者の頭文字で標識された)を横軸にプロットする。 (C)”キャプチャインデックス”は、ボトムアップの注意の尺度であり、タスクに無関係なディストラクターがタスクから注意を奪う程度を定量化します。 コントロールとSPL lesionedグループは、両方のグループがタスクに無関係なデストラクタによってキャプチャされるように、同様のキャプチャ値を示します。 TPJ病変群は捕獲指数が大幅に減少した(捕獲失敗)。 患者は、病変に基づくのではなく、行動に基づいてSPLまたはTPJグループに配置されたことに注意してください、したがって、患者が対応するグループに終わ
トップダウン選択とボトムアップ選択の相互作用
目標指向配向とSPLと刺激駆動配向とTPJの間には明らかに強い関連性があるが、Shomstein et al. (2010)patient studyは、これら2つのシステムが完全に独立しているわけではないことを示唆している。 この結論は、SPL損傷を有する患者が”ハイパーキャプチャ”としてラベル付けされた性能のパターンを示したという知見によって支持されている。”ターゲット着色されたdistractorだけが注意を捕獲した対照とは違って(ターゲット正確さを下げるために導く)、関係がない着色されたdistractorはまたSPLの損害の患者のために気が散ることを証明した。 さらに、対照のために注意はターゲットの手始めに先行したときだけdistractorsによって捕獲されたのに対し、SPLの損害の患者のために注意はターゲットと同時に このパフォーマンスパターンは、次のフレームワークで説明できます: SPLは、タスクに関連する刺激の側面を決定することを含む注意のトップダウンガイダンスを担当する(例えば、search for red target;Corbetta and Shulman,2 0 0 2;Serences e t a l., 2005). 次に、この注意セットは、TPJによって媒介される注意メカニズムの捕捉が、タスク関連情報(例えば、注意を捕捉するred distractors、およびred targetを検索するときに注意を捕捉しないgray distractors)によってのみ誘発されるように、TPJを制約する。 SPLの不在は、タスク関連の注意セットの確立を防ぎ、したがって、タスク関連の有無にかかわらず、任意の刺激が重要であるとみなされるため、注意(例えば、splグループのための注意をキャプチャするタスク無関係なディスストラクタ)を無差別にキャプチャする。
SPLとTPJは、少なくとも二つの可能な方法のいずれかで相互作用することが示唆されています。 第一の可能性は、TPJが行動関連刺激を検出するが、高い空間分解能を欠いている警告システムとして機能することであり、したがって、行動関連刺激が検出されたとき、その正確な位置は空間マップを格納するSPLによって供給される(Kastner et al. ら、1 9 9 9;WojciulikおよびKanwisher、1 9 9 9;BisleyおよびGoldberg、2 0 0 3;Silver e t a l., 2005). 関連する仮説的可能性は、行動的に関連する刺激が提示されたときに、捕捉機構(TPJを含む)が進行中の認知活動の遮断器として作用することである(Corbetta and Shulman、2002、2011)。 保存されたTPJを有するが病変SPLを有する患者で観察される活動の「超捕捉」パターンは、TPJが手元のタスクを終了させる制御信号を発し、それによって回路遮断器となると見なすという仮説のさらなる証拠を提供する(Corbetta and Shulman,2 0 0 2;Serences e t a l., 2005). 様々なパラダイムを用いた他の最近の神経イメージング研究は、トップダウンとボトムアップの注意指向の間の対話的な関係のためのさらなる証拠を提 ら、2 0 0 5;Asplund e t a l., 2010; とができる。
二つの注意系と二つの注意基質(SPLとTPJ)との間の相互作用の証拠は強いが、この相互作用がSPLとTPJとの間で直接であるか、または他の中間領域を介して達成されているかどうかは不明である。 先に述べたように、トップダウンとボトムアップの注意指向ネットワークは前頭葉内の様々な領域に関与しているため、二つのシステム間の収束は前頭葉を介して達成される可能性があると仮定することは合理的である。 トップダウンとボトムアップの注意選択の間の相互作用を調査している二つの最近の研究は、刺激駆動型と目標指向型選択の間の収束の可能性のある部位としてIFJとIFGの証拠を提供した(Asplund et al. ら,2 0 1 0;DiquattroおよびGeng,2 0 1 1)。 IFJおよびIFGは、注意および認知制御への一般的な関与、ならびに空間的および非空間的選択の両方への関与を考えると、そのような相互作用の理想的な候補であると思われる(Koechlin et al. ら、2 0 0 3;Bruss e t a l., 2005).トップダウンとボトムアップの選択における頭頂葉の役割
: Physiology
このレビューの重点は主に人間の研究に置かれてきましたが、注意指向における頭頂皮質の関与に関する豊富な知識は、サルの生理学調査から収集されています(bisley And Goldberg,2010による最近のレビューを参照してください)。 しかし、頭頂皮質内の異なる領域のトップダウンおよびボトムアップの注意指向への相対的な寄与を調べることになると、サル生理学の文献は短 これの主な理由は、サルの皮質内には、トップダウンとボトムアップのコントロールの同じ分離の証拠がないように見えることです。 代わりに、もともとサッカード計画に関与していると考えられている側頭頂内領域(LIP)(Gnadt and Andersen、1988)は、視覚的注意に関与し、外部刺激がトップダウンまたはボトムア, 2006; Buschman and Miller,2007;Gottlieb and Balan,2010).
結論
ヒトの頭頂皮質については、相同サル皮質よりもはるかに知られていないが、神経画像および神経心理学的方法を用いた最近の研究では、ますますきめ細かな機能的および構造的区別を解明し始めている。 トップダウンとボトムアップの注意指向における頭頂皮質の背側および腹側領域の認知的役割を解明する最近の神経イメージングおよび神経心理学的研究と、二つの注意割り当てメカニズムの間の相互作用に焦点を当てた。
利益相反に関する声明
著者は、潜在的な利益相反として解釈される可能性のある商業的または財務的関係がない場合に研究が行われた
謝辞
この作業は、国立衛生研究所の助成金EY021644によって支援されました。
Asplund,C.L.,Todd,J.J.,Snyder,A.P.,And Marois,R.(2010). 目標指向および刺激駆動の注意における外側前頭前野の中心的な役割。 ナット ニューロシ… 13, 507–512.
Pubmed Abstract|Pubmed Full Text/CrossRef Full Text
ベーコン、W.F.、およびEgeth、H.E.(1994)。 刺激駆動の注意捕獲をオーバーライドします。 パーセプト サイコフィーズ 55, 485–496.
Pubmed要約|Pubmed全文
バラン、P.F. およびGottlieb,J.(2 0 0 6)。 注意を撹乱することによって明らかにされた動的salienceマップへの外因性入力の統合。 J.Neurosci. 26, 9239–9249.
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Behrmann,M.,Geng,J.J.,And Shomstein,S.(2004). 頭頂皮質と注意。 カー オピン ニューロビオール 14, 212–217.
Pubmed Abstract|Pubmed Full Text/CrossRef Full Text
Bisley,J.W.,And Goldberg,M.E.(2003). 外側頭頂内領域における神経活動および空間的注意。 科学299,81-86.
Pubmed Abstract|Pubmed Full Text/CrossRef Full Text
Bisley,J.W.,And Goldberg,M.E.(2010). 頭頂葉における注意、意図、および優先順位。 アンヌ ニューロシー牧師。 33, 1–21.
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Brass,M.,Derrfuss,J.,Forstmann,B.,and von Cramon,D.Y.(2005). 認知制御における下前頭接合領域の役割。 トレンドCogn。 サイ… 9, 314–316.
Pubmed Abstract|Pubmed Full Text/CrossRef Full Text
Buschman,T.J.,And Miller,E.K.(2007). 前頭前野および後頭頂皮質における注意のトップダウン対ボトムアップ制御。 科学315、1860-1862。
Pubmed Abstract|Pubmed Full Text/CrossRef Full Text
Chiu,Y.C.,And Yantis,S.(2009). タスクセットの認知制御のドメインに依存しないソース:空間的な注意のシフトと分類ルールの切り替え。 J.Neurosci. 29, 3930–3938.
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Colby,C.L.,And Goldberg,M.E.(1999). 頭頂皮質における空間と注意。 アンヌ ニューロシー牧師。 22, 319–349.
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Corbetta,M.,Kincade,J.M.,Ollinger,J.M.,McAvoy,M.P.,And Shulman,G.L.(2000). 自発的な配向は、ヒト後頭頂皮質における標的検出から解離する。 ナット ニューロシ… 3, 292–297.
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Corbetta,M.,Kincade,M.J.,Lewis,C.,Snyder,A.Z.,And Sapir,A.(2005). 空間的無視における空間的注意欠陥の神経基盤と回復。 ナット ニューロシ… 8, 1603–1610.
Pubmed Abstract|Pubmed Full Text/CrossRef Full Text
Corbetta,M.,Patel,G.,And Shulman,G.L.(2008). 人間の脳の再配向システム:環境から心の理論へ。 58,306-324
Pubmed Abstract|Pubmed Full Text/CrossRef Full Text
Corbetta,M.,And Shulman,G.L.(2002). 脳内の目標指向および刺激駆動の注意の制御。 ナット ニューロシー牧師。 3, 201–215.
Pubmed Abstract|Pubmed Full Text/CrossRef Full Text
Corbetta,M.,And Shulman,G.L.(2011). 空間ネグレクトと注意ネットワーク。 アンヌ ニューロシー牧師。 34, 569–599.
Pubmed Abstract|Pubmed Full Text/CrossRef Full Text
Critchley,M.(1953). 頭頂葉。 イギリス・ロンドン:Hafner Press。pubmed Abstract/Pubmed Full Text
Diquattro,N.E.,And Geng,J.J.(2011). 文脈的知識は、注意制御ネットワークを構成する。 J.Neurosci. 31, 18026–18035.
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Downar,J.,Crawley,A.P.,Mikulis,D.J.,And Davis,K.D.(2001). 視覚および聴覚刺激の変化に対する皮質応答に対するタスク関連性の影響:イベント関連fMRI研究。 1256年-1267年。
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Downar,J.,Crawley,A.P.,Mikulis,D.J.,And Davis,K.D.(2002). 複数の感覚モダリティ間で中立的な行動のコンテキストで刺激salienceに敏感な皮質ネットワーク。 J.Neurophysiol。 87, 615–620.pubmed Abstract/Pubmed Full Text
Drummond,L.,And Shomstein,S.(2010). オブジェクトベースの注意: シフトまたは不確実性? アテン パーセプト サイコフィーズ 72, 1743–1755.
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Egeth,H.E.,And Yantis,S.(1997). 視覚的な注意:制御、表現、および時間コース。 アンヌ サイコル牧師 48, 269–297.
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Eriksen,C.W.,And Hoffman,J.E.(1972). 視覚表示からの選択的符号化の時間的および空間的特性。 パーセプト サイコフィーズ 12, 201–204.
Folk,C.L.,Leber,A.B.,Egeth,H.E.(2002). あなたが点滅しました! 偶発的な注意キャプチャは、空間的な瞬きを生成します。 パーセプト サイコフィーズ 64, 741–753.pubmed Abstract/Pubmed Full Text
Folk,C.L.,Remington,R.W.,And Johnston,J.C.(1992). 不随意の秘密の方向づけは注意制御設定によって偶発的である。 J.Exp. サイコル ハム パーセプト 実行します。 18, 1030–1044.Pubmed Abstract|Pubmed Full Text
Friedrich,F.J.,Egly,R.,Rafal,R.D.,And Beck,D.(1998). ヒトにおける空間的注意欠陥:上頭頂部病変と側頭頂部接合部病変の比較。 神経心理学12,193-207.pubmed Abstract|Pubmed Full Text
Geng,J.J.,And Behrmann,M.(2002). ターゲット位置の確率のcuingはhemispatial無視の正常な関係者そして患者の視覚調査を暗黙に促進する。 サイコル サイ… 13, 520–525.pubmed Abstract|Pubmed Full Text
Geng,J.J.,And Behrmann,M.(2005). 視覚的な検索における注意手がかりとしての空間的確率。 パーセプト サイコフィーズ 67, 1252–1268.pubmed Abstract|Pubmed Full Text
Geng,J.J.,And Mangun,G.R.(2011). 顕著な文脈キューによる右側頭頭頂接合活性化は、ターゲット識別を容易にする。 54,594-601pubmed Abstract/Pubmed Full Text|CrossRef Full Text
Gibson,B.S.,And Kelsey,E.M.(1998). 刺激主導の注意の捕獲はdisplaywideの視覚特徴のための注意セットに偶発的である。 J.Exp. サイコル ハム パーセプト 実行します。 24, 699–706.
Pubmed Abstract|Pubmed Full Text
Giesbrecht,B.,Woldorff,M.G.,Song,A.W.,And Mangun,G.R.(2003). 空間的および特徴的な注意の間のトップダウン制御の神経機構。 19,496-512
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Gnadt,J.W.,And Andersen,R.A.(1988). マカクの後頭頂皮質における記憶関連運動計画活動。 経験値 脳Res.70,216-220.pubmed Abstract/Pubmed Full Text
Gottlieb,J.,And Balan,P.(2010). 情報空間における決定としての注意。 トレンドCogn。 サイ… 14, 240–248.
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Greenberg,A.S.,Esterman,M.,Wilson,D.,Serences,J.T.,And Yantis,S.(2010). 前頭頂皮質における空間的および特徴ベースの注意の制御。 J.Neurosci. 30, 14330–14339.
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Hopfinger,J.B.,Buonocore,M.H.,And Mangun,G.R.(2000). トップダウンの注意制御の神経機構。 ナット ニューロシ… 3, 284–291.
Pubmed要約|Pubmed全文|CrossRef全文
Ipata,A.E. ることができると考えられています。 ポップアウト刺激に対する唇応答は、それが明白に無視される場合に減少する。 ナット ニューロシ… 9, 1071–1076.pubmed Abstract/Pubmed Full Text/CrossRef Full Text
Johnston,W.A.,And Dark,V.J.(1986). 選択的な注意。 アンヌ サイコル牧師 37, 43–75.
Juola,J.F.,Koshino,H.,およびWarner,C.B.(1995). 空間的な手がかりの注意効果と突然のオンセットとの間のトレードオフ。 パーセプト サイコフィーズ 57, 333–342.pubmed Abstract|Pubmed Full Text
Kastner,S.,Pinsk,M.A.,De Weerd,P.,Desimone,R.,And Ungerleider,L.G.(1999). 視覚刺激の非存在下での指示された注意の間の人間の視覚野における活動の増加。 22,751-761
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Koechlin,E.,Ody,C.,And Kouneiher,F.(2003). 人間の前頭前野における認知制御のアーキテクチャ。 科学302、1181-1185。
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Koshino,H. およびJuola,J. 視覚的注意における中枢、末梢、および突然の手掛かりの相対的な有効性。 Q.J.Exp. サイコル 45,609-631
Pubmed Abstract|Pubmed Full Text
Liu,T.,Slotnick,S.D.,Serences,J.T.,And Yantis,S.(2003). 特徴ベースの注意制御の皮質機構。 セレブ 1334年-1343年。
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Marois,R.,Leung,H.C.,And Gore,J.C.(2000). 人間の脳におけるオブジェクトの同一性と位置処理への刺激駆動型のアプローチ。 25,717-728
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
McFire,J.,And Zangwill,O.L.(1960). Visuo-左大脳半球の病変に関連する建設的な障害。 脳82,243-260.
特徴ベースの注意は視覚処理にどのように影響しますか? J.Exp. サイコル ハム パーセプト 実行します。 24, 1296–1310.
div およびDesimone,R. 選択的注意は、筋外皮質における視覚処理をゲートします。 科学229,782-784.
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
O’Craven,K.,Rosen,B.R.,Kwong,K.K.,Treisman,A.,And Savoy,R.L.(1997). 自発的な注意は、ヒトMT-MSTにおけるfMRI活性を調節する。 18,591-598pubmed Abstract/Pubmed Full Text|CrossRef Full Text
Piercy,M.(1964). 脳病変が知的機能に及ぼす影響:現在の研究動向のレビュー。 Br. J.精神医学110、310-352。pubmed Abstract/Pubmed Full Text
Posner,M.I.(1980). 注意の方向づけ。 Q.J.Exp. サイコル 32, 3–25.pubmed Abstract|Pubmed Full Text
Posner,M.I.,Snyder,C.R.,And Davidson,B.J.(1980). 注意と信号の検出。 J.Exp. サイコル 109, 160–174.pubmed Abstract|Pubmed Full Text
Posner,M.I.,Walker,J.A.,Friedrich,F.J.,And Rafal,R.D.(1984). 頭頂部損傷が注意の秘密の方向付けに及ぼす影響。 J.Neurosci. 4, 1863–1874.
Pubmed Abstract|Pubmed Full Text
Serences,J.T.,Shomstein,S.,Leber,A.B.,Golay,X.,Egeth,H.E.,And Yantis,S.(2005). 人間の皮質における自発的および刺激駆動型の注意制御の調整。 サイコル サイ… 16, 114–122.
Pubmed Abstract|Pubmed Full Text/CrossRef Full Text
Shaw,M.L.(1978). 反応時間の容量割り当てモデル。 J.Exp. サイコル ハム パーセプト 実行します。 4, 586–598.
オブジェクトと空間的な場所の両方に視覚的な注意を仲介する皮質システム。 プロク… ナトル アカド サイ… 103,11387-11392.
Pubmed Abstract|Pubmed Full Text/CrossRef Full Text
Shomstein,S.,Lee,J.,And Behrmann,M.(2010). トップダウンおよびボトムアップの注意指導:背側および腹側頭頂皮質の役割を調査する。 経験値 脳Res.206,197-208.
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Shomstein,S.,And Yantis,S.(2004a). オブジェクトベースの注意における構成とコンテキストの優先順位付け。 サイコン ブル 11,247-253.pubmed Abstract/Pubmed Full Text
Shomstein,S.,And Yantis,S.(2004b). 人間の皮質における視覚と聴覚の間の注意シフトの制御。 J.Neurosci. 24, 10702–10706.
Pubmed Abstract|Pubmed Full Text/CrossRef Full Text
Shomstein,S.,And Yantis,S.(2006). 頭頂皮質は、空間的および非空間的な聴覚注意の自発的な制御を仲介する。 J.Neurosci. 26, 435–439.
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
シルバー、M.A.、Ress、D。 およびHeeger,D.J.(2 0 0 5)。 人間の頭頂皮質における視覚空間的注意の地形図。 J.Neurophysiol。 94, 1358–1371.
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Tamber-Rosenau,B.J.,Esterman,M.,Chiu,Y.-C.,And Yantis,S.(2011). 視覚と作業記憶における注意のシフトのための認知制御の皮質メカニズム。 J.Cogn. ニューロシ… 23, 2905–2919.
Pubmed Abstract|Pubmed Full Text/CrossRef Full Text
Theeuwes,J.(1991). 注意の外因性および内因性制御: 視覚的なオンセットとオフセットの効果。 パーセプト サイコフィーズ 49, 83–90.pubmed Abstract/Pubmed Full Text
Vallar,G.,And Perani,D.(1986). 右半球の打撃の損害の後の一方的な無視の解剖学。 ヒトにおける臨床/CTスキャン相関研究。 神経心理学24、609-622。
Pubmed Abstract|Pubmed Full Text/CrossRef Full Text
Wojciulik,E.,And Kanwisher,N.(1999). 視覚的注意における頭頂部の関与の一般性。 23,747-764
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Yantis,S.,And Jonides,J.(1984). 突然の視覚的な不調と選択的注意:視覚的検索からの証拠。 J.Exp. サイコル ハム パーセプト 実行します。 10, 601–621.pubmed Abstract/Pubmed Full Text
Yantis,S.,And Jonides,J.(1990). 突然の視覚的なオンセットと選択的な注意:自発的な割り当てと自動割り当て。 J.Exp. サイコル ハム パーセプト 実行します。 16, 121–134.pubmed Abstract|Pubmed Full Text
Yantis,S.,Schwarzbach,J.,Serences,J.T. とができるようになることを期待しています。 空間的注意シフト中のヒト頭頂皮質における一過性の神経活動。 ナット ニューロシ… 5, 995–1002.
Pubmed Abstract|Pubmed Full Text|CrossRef Full Text
Yantis,S.,And Serences,J.T.(2003). 空間ベースおよびオブジェクトベースの注意制御の皮質メカニズム。 カー オピン ニューロビオール 13, 187–193.
Pubmed要約|Pubmed全文|CrossRef全文