学習目標
このセクションの終わりまでに、あなたは次のことができます:
- 解糖のプロセスを説明
- クレブスサイクルを介してピルビン酸分子の経路を説明
- 電子輸送鎖を介して電子の輸送を説明
- 酸化的リン酸化 炭水化物のファミリーには、単純な糖と複雑な糖の両方が含まれています。 グルコースとフルクトースは単純糖の例であり、デンプン、グリコーゲン、セルロースはすべて複雑な糖の例である。 複合糖は多糖類とも呼ばれ、複数の単糖類分子でできています。 多糖類は、エネルギー貯蔵(例えば、デンプンおよびグリコーゲン)として、および構造成分(例えば、昆虫のキチンおよび植物のセルロース)として役立つ。
消化中、炭水化物は単純な可溶性糖に分解され、腸壁を越えて循環系に輸送され、体全体に輸送されます。
消化中、炭水化物は腸壁を越えて循環系に輸送され、体全体に輸送されます。
水化物の消化はでんぷんの唾液のアミラーゼの行為の口で始まり、小腸の上皮を渡って吸収される単糖類で終わります。 吸収された単糖が組織に輸送されると、細胞呼吸のプロセスが始まります(図1)。 このセクションでは、最初に解糖、単糖グルコースが酸化され、その結合に蓄積されたエネルギーを放出してATPを生成するプロセスに焦点を当てます。
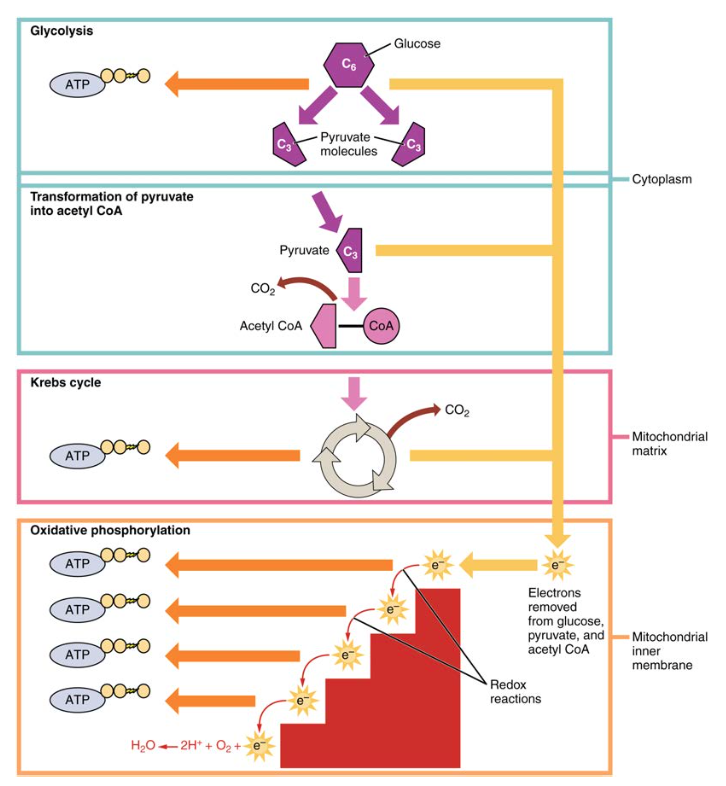
図1. 細胞呼吸は解糖、Krebs周期およびATPを作り出すために酸化的リン酸化によってブドウ糖の分子を酸化します。
解糖
グルコースは、身体の最も容易に利用可能なエネルギー源です。 消化プロセスが多糖類をグルコースを含む単糖類に分解した後、単糖類は小腸の壁を越えて循環系に輸送され、肝臓に輸送されます。 肝臓では、肝細胞は循環系を介してグルコースを通過させるか、または過剰なグルコースをグリコーゲンとして貯蔵する。 体内の細胞はインスリンに応答して循環グルコースを取り込み、解糖と呼ばれる一連の反応を通じて、グルコース中のエネルギーの一部をADPに移してATPを形成します(図2)。 解糖の最後のステップは、生成物ピルビン酸塩を生成する。
解糖は、ヘキソキナーゼによるグルコースのリン酸化から始まり、グルコース-6-リン酸を形成する。 このステップは、リン酸基の供与体である1つのATPを使用する。 ホスホフルクトキナーゼの作用下で、グルコース-6-リン酸はフルクトース-6-リン酸に変換される。 この時点で、第二のATPは、フルクトース-1,6-ビスリン酸を形成し、そのリン酸基を寄付します。 この六炭素糖は、グリセルアルデヒド-3-リン酸とジヒドロキシアセトンリン酸の二つのリン酸化三炭素分子を形成するために分割され、両方がグリセルアルデヒド-3-リン酸に変換される。 グリセルアルデヒド-3-リン酸は、細胞内に存在するリン酸二水素によって供与された基でさらにリン酸化され、三炭素分子1,3-ビスホスホグリセリン酸を形成する。 この反応のエネルギーは、グリセルアルデヒド-3-リン酸の酸化(からの電子の除去)に由来する。 ピルビン酸に至る一連の反応では、2つのリン酸基が2つのAdpに移動して2つのAtpを形成する。 したがって、解糖は2つのAtpを使用するが、4つのAtpを生成し、2つのAtpと2つのピルビン酸分子の純利得をもたらす。 酸素の前で、ピルビン酸塩はKrebs周期に続きます(また付加的なエネルギーが得られ、渡されるクエン酸周期かtricarboxylic酸周期(TCA)と呼ばれる)。
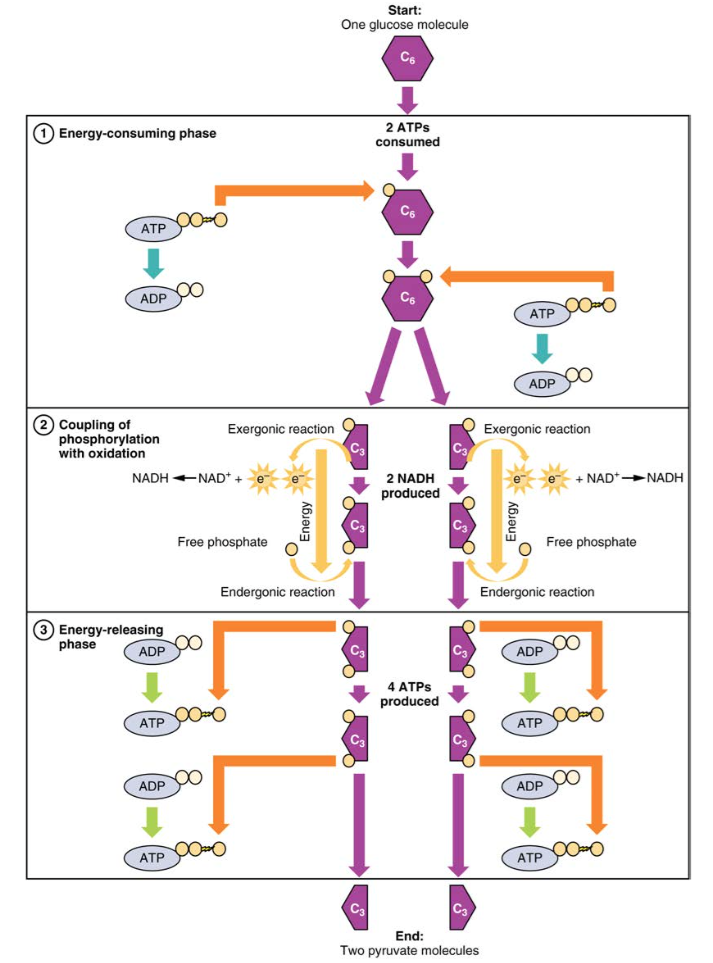
図2。 解糖のエネルギー消費段階では、2つのAtpが消費され、2つのリン酸塩がグルコース分子に移動します。 その後、グルコース分子は2つの3炭素化合物に分割され、それぞれにリン酸塩が含まれています。 第二相の間に、追加のリン酸塩が三炭素化合物のそれぞれに添加される。 このエンダーゴン反応のエネルギーは、それぞれの3炭素化合物から2つの電子を除去(酸化)することによって提供される。 エネルギー放出相の間に、リン酸塩は両方の3炭素化合物から除去され、4つのATP分子を生成するために使用される。解糖について学ぶためにこのビデオを見てください:
解糖は、エネルギー消費(化学プライミングとも呼ばれる)とエネルギー降伏の二つの相に分けるこ 最初の相はエネルギー消費相であるため、グルコースの各分子に対して反応を開始するには2つのATP分子が必要です。 しかし、反応の終わりには4つのAtpが生成され、その結果、2つのATPエネルギー分子の正味の利得が得られる。解糖は、次の式で表すことができます。
解糖は、次の式で表すことができます。:
\text{Glucose}+2\text{ATP}+2\text{NAD}^{+}+4\text{ADP}+2\text{P}_{i}\to\text{Pyruvate}+4\text{ATP}+2\text{NADH}+2\text{H}^{+}
この式は、グルコースがATP(エネルギー源)、NAD+(電子受容体として機能する補酵素)、無機リン酸と組み合わせて、二つのピルビン酸分子に分解し、二つのATP分子と二つのエ このプロセスで生産されるNADHは、後でミトコンドリアでATPを生産するために使用されます。 重要なことに、このプロセスの終わりまでに、1つのグルコース分子は2つのピルビン酸分子、2つの高エネルギー ATP分子、および2つの電子を運ぶNADH分子を生成する。解糖の以下の議論には、反応の原因となる酵素が含まれる。
グルコースが細胞に入ると、酵素ヘキソキナーゼ(または肝臓のグルコキナーゼ)は急速にリン酸を加えてグルコース-6-リン酸に変換する。 キナーゼは、基質にリン酸分子を添加する酵素の一種である(この場合、グルコースであるが、他の分子にも当てはまる可能性がある)。 この変換ステップは、1つのATPを必要とし、本質的に細胞内のグルコースを捕捉し、それが原形質膜を通過するのを防ぎ、解糖が進行することを可能にする。 それはまたティッシュのより血の高いブドウ糖のレベルの集中の勾配を維持するために作用します。 この濃度勾配を確立することによって、血液中のグルコースは、高濃度の領域(血液)から、使用または貯蔵される低濃度の領域(組織)に流れることができる。 ヘキソキナーゼは、体内のほぼすべての組織に見られます。 グルコキナーゼは、一方では、血ブドウ糖のレベルが高いとき活動的であるレバーのようなティッシュで表現されます。 ヘキソキナーゼにglucokinaseよりブドウ糖のための高い類縁があり、従ってglucokinaseより速い率でブドウ糖を変えられます。 これは、グルコースがより多く必要とする組織に優先的に移動することを可能にするので、グルコースのレベルが体内で非常に低い場合に重要である。
解糖の第一段階の次のステップでは、酵素グルコース-6-リン酸イソメラーゼはグルコース-6-リン酸をフルクトース-6-リン酸に変換する。 ブドウ糖のように、フルクトースはまた6つのカーボン含んでいる砂糖です。 酵素のphosphofructokinase-1はそれからフルクトースにフルクトース6隣酸塩を変えるためにもう一つの隣酸塩を加えます-1-6-ビスリン酸、別のATP分子を使用して、別の六炭素糖、。 アルドラーゼはこのフルクトースを分解し-1-6-二つの三炭素分子、グリセルアルデヒド-3-リン酸とジヒドロキシアセトンリン酸にビスリン酸。 トリオセリン酸イソメラーゼは、ジヒドロキシアセトンリン酸を第二のグリセルアルデヒド-3-リン酸分子に変換する。 従って、この化学プライミングかエネルギー消費段階の終わりまでに、1つのブドウ糖の分子は2つのglyceraldehyde-3隣酸塩の分子に破壊されます。解糖の第二段階、エネルギー降伏段階は、解糖の産物であるエネルギーを生成する。
解糖の第二段階は、解糖の生成物であるエネルギーを生成する。
解 グリセルアルデヒド-3-リン酸デヒドロゲナーゼは、
エネルギー消費相中に生成された各三炭素グリセルアルデヒド-3-リン酸を1,3-ビスホスホグリセリン酸に変換する。 この反応は電子を放出し、それがNAD+によって拾われてNADH分子を生成する。 NADHはATPのような高エネルギー分子ですが、ATPとは異なり、細胞によってエネルギー通貨として使用されません。 2つのグリセルアルデヒド-3-リン酸分子が存在するので、2つのNADH分子がこのステップの間に合成される。 各1,3-ビスホスホグリセリン酸は、その後、ホスホグリセリン酸キナーゼによって3-ホスホグリセリン酸に脱リン酸化される(すなわち、リン酸が除去される)。 この反応で放出された各リン酸塩は、ADPの1分子を1つの高エネルギー ATP分子に変換し、2つのATP分子を得ることができる。
酵素ホスホグリセリン酸ムターゼは、3-ホスホグリセリン酸分子を2-ホスホグリセリン酸に変換する。 その後、エノラーゼ酵素は2-ホスホグリセリン酸分子に作用してホスホエノールピルビン酸分子に変換する。 解糖の最後のステップは、ピルビン酸キナーゼによる2つのホスホエノールピルビン酸分子の脱リン酸化を伴い、2つのピルビン酸分子と2つのATP分子を生成する。要約すると、1つのグルコース分子が2つのピルビン酸分子に分解され、解糖によって2つの正味のATP分子と2つのNADH分子が生成されます。
従って、解糖は細胞のためのエネルギーを発生させ、好気性のKrebs周期(またクエン酸周期かtricarboxylic酸周期と呼ばれる)によって更に処理することができるpyruvateの分子を作成します;発酵によって乳酸かアルコールに(イーストで)変えられて;またはgluconeogenesisによってブドウ糖の統合のために後で使用されます。酸素が限られているか存在しない場合、ピルビン酸塩は嫌気性経路に入る。
嫌気性呼吸
酸素が限られているか存在しない場合、ピルビン酸塩は嫌気性経路に入る。
嫌気性呼吸れらの反応では、ピルビン酸は乳酸に変換することができる。 追加のATPを生成することに加えて、この経路はピルビン酸濃度を低く保つのに役立ち、解糖が継続し、解糖によって必要とされるNAD+にNADHを酸化する。 この反応では、乳酸は最終的な電子受容体として酸素を置換する。 嫌気性呼吸は、酸素が限られているか、またはミトコンドリアが存在しないか、または機能していないときに、身体のほとんどの細胞で発生する。 例えば、赤血球(赤血球)はミトコンドリアを欠いているため、嫌気性呼吸からATPを産生しなければならない。 これは、数秒から数分の範囲で、短時間のATP産生の効果的な経路である。 生成された乳酸は血漿中に拡散し、肝臓に運ばれ、そこでcoriサイクルを介してピルビン酸またはグルコースに変換される。 同様に、人が運動するとき、筋肉は酸素がそれらに送達されることができるよりも速くATPを使用する。 それらは、迅速なATP産生のための解糖および乳酸産生に依存する。
好気性呼吸
酸素の存在下では、ピルビン酸塩はクレブスサイクルに入ることができ、電子がピルビン酸塩から受容体NAD+、GDP、FADに移動し、二酸化炭素が”廃棄物”となるように追加のエネルギーが抽出される(図3)。 NADHおよびFADH2はATPを作り出すのに移動されたエネルギーを使用する電子輸送鎖に電子を渡します。 電子輸送鎖の末端ステップとして、酸素は末端電子受容体であり、ミトコンドリア内に水を生成する。
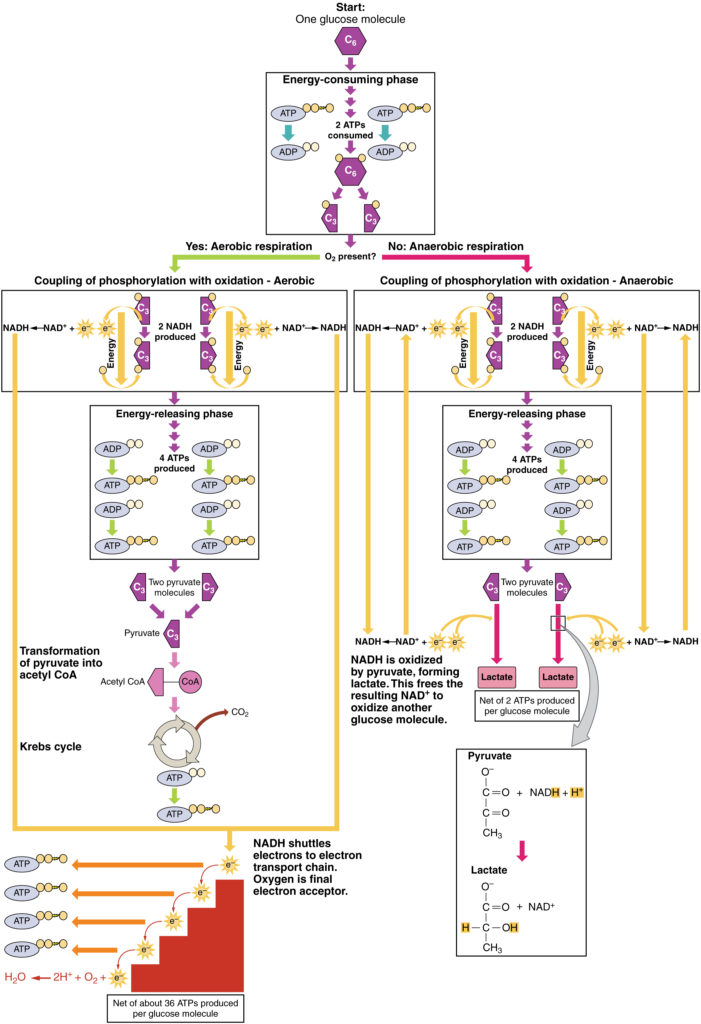
図3. クリックすると拡大画像が表示されます。 嫌気性呼吸のプロセスは、酸素の非存在下で、またはミトコンドリアを欠いている赤血球内でグルコースを二つの乳酸分子に変換する。 好気性呼吸の間に、グルコースは2つのピルビン酸分子に酸化される。
Krebsサイクル/クエン酸サイクル/トリカルボン酸サイクル
解糖中に生成されたピルビン酸分子は、ミトコンドリア膜を横切ってミトコンドリア内マトリックスに輸送され、そこでKrebsサイクルと呼ばれる経路の酵素によって代謝される(図4)。 クレブスサイクルは、一般的にクエン酸サイクルまたはトリカルボン酸(TCA)サイクルとも呼ばれます。 クレブス周期の間に、ATP、NADHおよびFADH2を含む高エネルギー分子は、作成されます。 NADHとFADH2は、ミトコンドリア内の電子輸送鎖を介して電子を通過させ、より多くのATP分子を生成する。
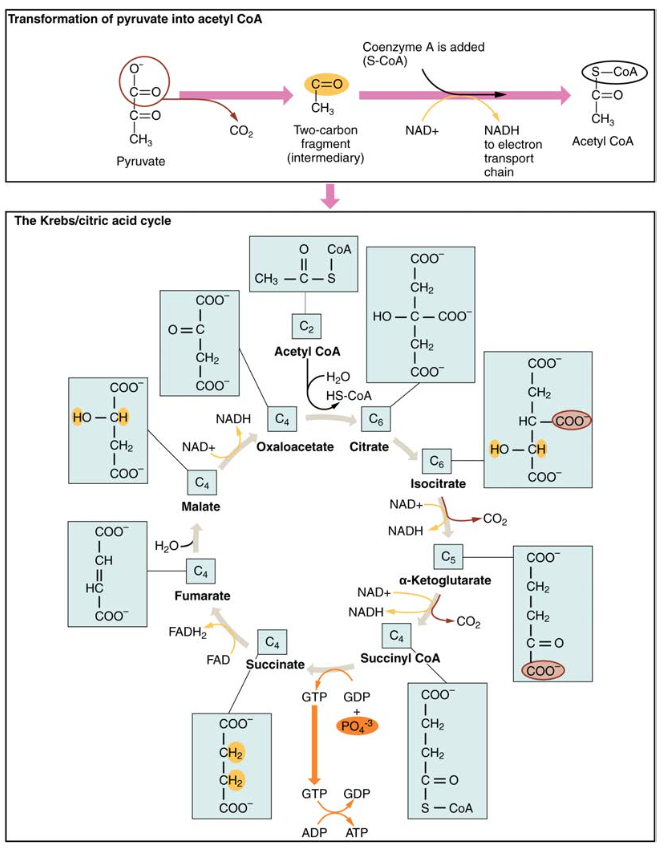
図4。 クレブス周期の間に、解糖によって発生する各ピルビン酸塩は二炭素アセチルCoAの分子に変えられます。 アセチルCoAは周期によって組織的に処理され、高エネルギー NADH、FADH2およびATPの分子を作り出します。
このアニメーションを見て、クレブス周期を観察してください。解糖中に生成された三炭素ピルビン酸分子は、細胞質からミトコンドリアマトリックスに移動し、そこで酵素ピルビン酸デヒドロゲナーゼによって二炭素アセチル補酵素A(アセチルCoA)分子に変換される。 この反応は酸化的脱炭酸反応である。 これは、三炭素ピルビン酸塩を二炭素アセチルCoA分子に変換し、二酸化炭素を放出し、NAD+と結合してNADHを形成する二つの電子を移動させる。 アセチルCoAは、四炭素分子、オキサロ酢酸と組み合わせることによってクレブスサイクルに入り、六炭素分子クエン酸またはクエン酸を形成し、同時に補酵素A分子を放出する。
六炭素クエン酸分子は体系的に五炭素分子に変換され、次に四炭素分子に変換され、サイクルの始まりであるオキサロ酢酸で終わる。 途中で、各クエン酸分子は1つのATP、1つのFADH2、および3つのNADHを生成します。 FADH2およびNADHは内部のmitochondrial膜にある酸化リン酸化システムに入ります。 さらに、Krebs周期は蛋白質および脂肪を処理し、破壊するために出発材料を供給する。
クレブス周期を始めるためには、クエン酸塩のシンターゼは六炭素クエン酸塩の分子を形作るためにアセチルCoAおよびoxaloacetateを結合します;CoAは続いて解放され、別のピルビン酸塩の分子と結合して周期を再度始めることができます。 アコニターゼ酵素はクエン酸塩をイソクエン酸塩に変換する。 酸化的脱炭酸の二つの連続したステップでは、イソシトレート脱水素酵素がイソシトレートを五炭素α-ケトグルタル酸に変換し、α-ケトグルタル酸デヒドロゲナーゼによって触媒され、四炭素スクシニルCoAに変換すると、CO2分子とNADH分子の二つの分子が生成される。 次に、スクシニルCoAデヒドロゲナーゼはスクシニルCoAをコハク酸に変換し、高エネルギー分子GTPを形成し、そのエネルギーをADPに移してATPを生成する。 コハク酸デヒドロゲナーゼは、コハク酸をフマル酸に変換し、FADH2の分子を形成する。 その後、フマル酸塩をリンゴ酸塩に変換し、リンゴ酸デヒドロゲナーゼはnad+をNADHに還元しながらオキサロ酢酸に変換する。 Oxaloacetateは次のアセチルCoAと結合して再度Krebs周期を始めるためにそれから準備ができています(図4を見て下さい)。 サイクルの各ターンについて、3つのNADH、1つのATP(GTPを介して)、および1つのFADH2が作成されます。 ピルビン酸塩の各炭素は、酸化的(好気性)呼吸の副産物として放出されるCO2に変換される。
酸化的リン酸化と電子輸送鎖
電子輸送鎖(ETC)は、クレブスサイクルによって生成されたNADHとFADH2を使用してATPを生成します。 NADHおよびFADH2からの電子は一連の酵素の反作用によって内部のmitochondrial膜で埋め込まれる蛋白質の複合体を通って移ります。 電子輸送鎖は、一連の4つの酵素複合体(複合体I–複合体IV)と2つの補酵素(ユビキノンとシトクロムc)で構成されており、これらは電子キャリアとプロトンポンプとして機能し、h+イオンをミトコンドリア内膜と外膜の間の空間に移動させます(図5)。 ETCは、ドナー(NADHのような)と電子受容体(O2のような)との間の電子の移動を、ミトコンドリア内膜を横切るプロトン(H+イオン)の移動と結合させ、酸化的リン酸化のプロセスを可能にする。 酸素の存在下では、エネルギーは電子キャリアを通って段階的に通過し、リン酸塩をADPに結合してATPを生成するのに必要なエネルギーを徐々に収集する。 分子酸素、O2の役割は等のための末端の電子受容器として、あります。 これは、電子がETC全体を通過すると、それらを別の別々の分子に渡さなければならないことを意味します。 マトリックスからのこれらの電子、O2、およびH+イオンは、新しい水分子を形成するために結合する。 これは酸素で呼吸するあなたの必要性のための基礎です。 酸素がなければ、ETCを通る電子の流れは止まる。
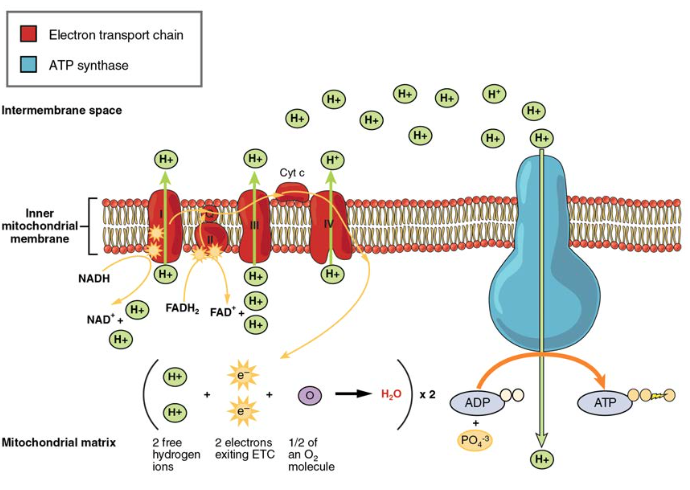
図5。 電子輸送鎖は、内部のミトコンドリアマトリックスからH+イオンを汲み出すために使用される一連の電子キャリアおよびイオンポンプである。電子輸送鎖について学ぶには、このビデオを見てください。
電子輸送鎖について学ぶには、このビデオを見てください。
ADHとFADH2から放出された電子は、それぞれのキャリアによって鎖に沿って渡され、電子を受け取ると還元され、次のキャリアに渡すと酸化されます。
これらの反応のそれぞれは、少量のエネルギーを放出し、H+イオンを内膜に送り出すために使用されます。 膜間の空間におけるこれらのプロトンの蓄積は、ミトコンドリアマトリックスに関してプロトン勾配を作成する。
また、ミトコンドリア内膜に埋め込まれているのは、ATPシンターゼと呼ばれる驚くべきタンパク質孔複合体です。 効果的には、勾配の下のそしてmitochondrialマトリックスへの内部の膜を渡るH+イオンの流れによって動力を与えられるのはタービンです。 H+イオンが複合体を横断すると、複合体の軸が回転します。 この回転により、ATP合成酵素の他の部分がADPおよびPiにATPを生成させることが可能になる。 好気性呼吸によってグルコース分子あたりに産生されるATPの総数を考慮する際には、以下の点を覚えておくことが重要です。
- 解糖によって産生されるATPのネット(エネルギー消費段階で産生されるものと消費されるものがある)。 しかし、これらの2つのATPは、解糖中に産生されたNADHを細胞質からミトコンドリアに輸送するために使用される。 したがって、解糖中のATPの正味産生はゼロである。解糖後のすべての段階で、生成されるATP、NADH、およびFADH2の数に2を乗算して、各グルコース分子が2つのピルビン酸分子をどのように生成するかを反映
- ETCでは、酸化されたNADHごとに約3つのATPが生成されます。 しかし、酸化されたFADH2ごとに約二つのATPのみが生成されます。 FADH2からの電子は、NADH(複合体I)からの電子と比較してETC(複合体II)の低い点で始まるため、ATPを生成しません(図5)。
したがって、好気性呼吸に入るすべてのグルコース分子に対して、正味の合計36のAtpが生成されます(図6参照)。
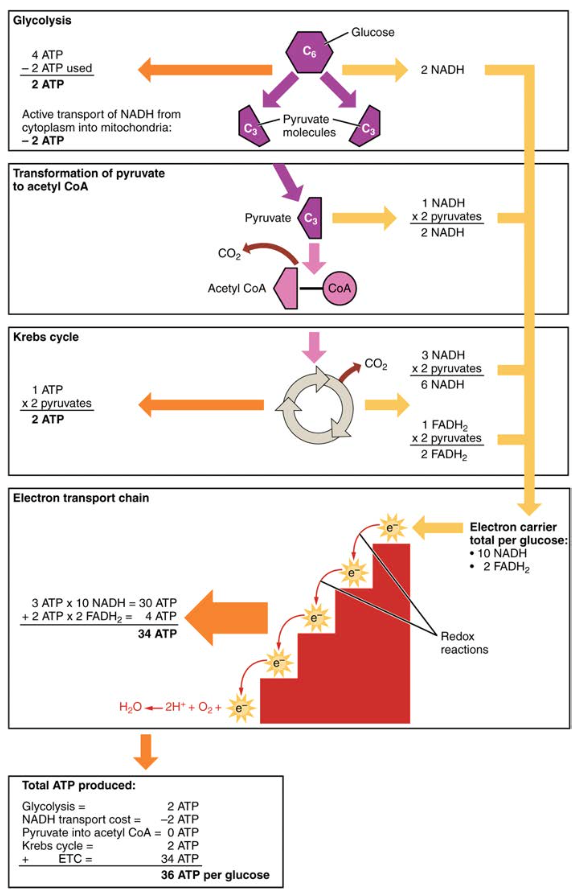
図6。 炭水化物の新陳代謝は解糖、Krebs周期および電子輸送の鎖を含みます。
糖新生
糖新生は、ピルビン酸塩、乳酸塩、グリセロール、またはアミノ酸アラニンまたはグルタミンからの新しいグルコース分子の合成で このプロセスは、主に低グルコースの期間、すなわち絶食、飢餓、および低炭水化物食の条件下で肝臓で行われる。 だから、なぜ体がそれだけで破壊するためにかなりの努力を費やしたものを作り出すのかという疑問を提起することができますか? 脳を含む特定の重要な器官は、エネルギー源としてグルコースのみを使用することができるため、体が最小の血糖濃度を維持することが不可欠である。 血ブドウ糖の集中がそのある特定のポイントの下で下るとき常態に血の集中を上げるために、新しいブドウ糖はレバーによって総合されます。
糖新生は単に解糖の逆ではありません。 いくつかの重要な違いがあります(図7)。 ピルビン酸塩は、糖新生のための一般的な出発物質である。 最初に、ピルビン酸塩はoxaloacetateに変えられます。 Oxaloacetateはphosphoenolpyruvate(PEP)にoxaloacetateを変形させる酵素のphosphoenolpyruvateのcarboxykinase(PEPCK)のための基質としてそれから役立ちます。 この段階から、糖新生は解糖のほぼ逆である。 PEPは2-ホスホグリセリン酸に変換され、3-ホスホグリセリン酸に変換される。 その後、3-ホスホグリセリン酸は1,3ビスホスホグリセリン酸に変換され、グリセルアルデヒド-3-リン酸に変換される。 グリセルアルデヒド-3-リン酸の二つの分子は、フルクトースを形成するために結合します-1-6-次にフルクトース6隣酸塩とブドウ糖6隣酸塩に変えられるbisphosphate。 最後に、一連の反応はグルコース自体を生成する。 糖新生(解糖と比較して)では、酵素ヘキソキナーゼはグルコース-6-ホスファターゼに置き換えられ、酵素ホスホフルクトキナーゼ-1はフルクトース-1,6-ビスホスファターゼに置き換えられる。 これは、細胞が互いに独立して解糖および糖新生を調節するのを助ける。脂肪分解の一部として議論されるように、脂肪はグリセロールに分解され、リン酸化されてジヒドロキシアセトンリン酸またはDHAPを形成することがで DHAPは、解糖経路に入るか、または糖新生のための基質として肝臓によって使用され得る。
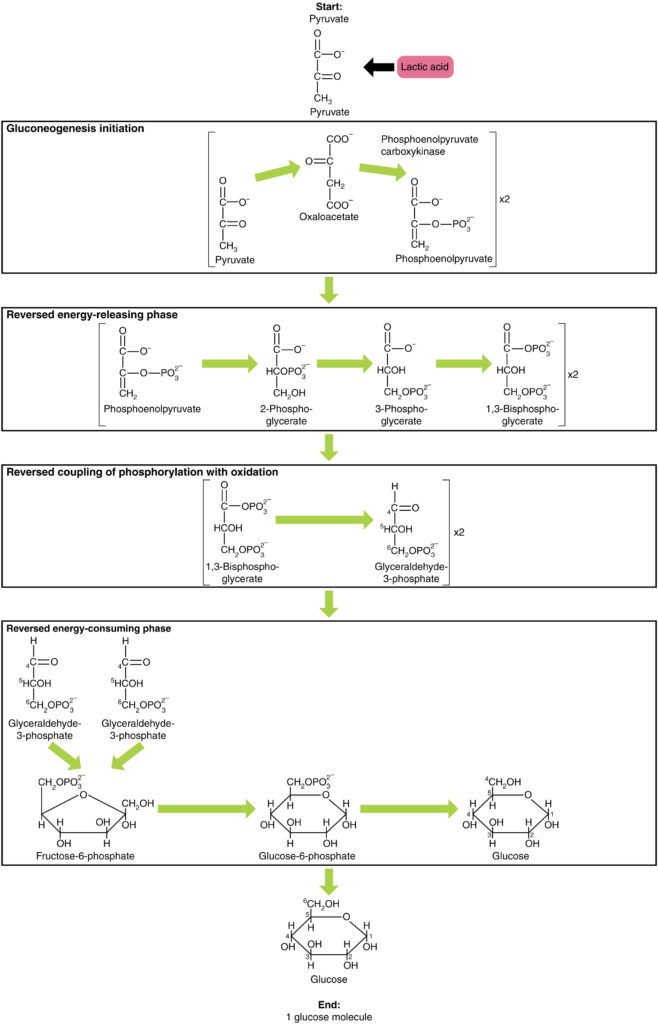
図7. クリックすると拡大画像が表示されます。 糖新生は、ピルビン酸、乳酸、グリセロール、アラニン、またはグルタミン酸からのグルコースの合成である。
老化と体の代謝率
人間の体の代謝率は、年齢30後十年あたりほぼ2パーセント減少します。 減らされた細い筋肉固まりを含むボディ構成の変更は、この減少に大抵責任があります。 筋肉固まりの最も劇的な損失、および代謝率の必然的な低下は50そして70年齢の間に、起こります。 筋肉量の損失は、高齢者が十分な身体活動に従事するのを阻害する傾向がある強度の低下と同等である。 これは減らされた身体活動がさらに筋肉損失をもたらす肯定的フィードバックシステムで起因し、更に新陳代謝を減らします。
代謝の一般的な低下を防ぎ、これらの低下の循環的性質と戦うために行うことができるいくつかのことがあります。
代謝の一般的な低下を防 これらは朝食を食べ、小さい食事を頻繁に食べ、沢山の細い蛋白質を消費し、水和させて残るために水を飲み、運動し(を含む強さの訓練)、および十分な睡眠を得ていることを含んでいる。 これらの措置は、エネルギーレベルが低下するのを防ぎ、過度の間食からのカロリー消費の増加の衝動を抑制するのに役立ちます。 これらの戦略は代謝を維持することは保証されていませんが、筋肉の損失を防ぐのに役立ち、エネルギーレベルを上げる可能性があります。 一部の専門家はまた、余分な脂肪貯蔵につながる可能性のある砂糖を避けることを示唆しています。 辛い食べ物や緑茶も有益かもしれません。 圧力がコルチゾール解放を活動化させ、コルチゾールが新陳代謝を遅らせるので、圧力を避けるか、または少なくとも弛緩の技術を練習して、また助けるこ
代謝酵素は、食物に含まれる炭水化物を分解する異化反応を触媒します。
代謝酵素は、食物に含まれる炭水化物を分解する異化反応を触媒します。
放出されたエネルギーは、あなたの体を構成する細胞やシステムに電力を供給するために使用されます。 過剰または未利用のエネルギーは、後の使用のために脂肪またはグリコーゲンとして保存される。 炭水化物代謝は口の中で始まり、そこで酵素唾液アミラーゼは複雑な糖を単糖に分解し始める。 これらは腸の膜を渡って血流にそしてボディティッシュにそれから運ぶことができます。 細胞内では、六炭素糖であるグルコースが一連の反応によってより小さな糖に処理され、分子内に貯蔵されたエネルギーが放出される。 炭水化物異化の最初のステップは解糖であり、これはピルビン酸、NADHおよびATPを産生する。 嫌気性条件下では、ピルビン酸塩を乳酸塩に変換して解糖を維持することができる。 好気性条件下では、ピルビン酸はクエン酸サイクルまたはトリカルボン酸サイクルとも呼ばれるクレブスサイクルに入る。 ATPに加えて、Krebsサイクルは、より高エネルギーのATP分子を生成する酸化的リン酸化プロセスに電子を提供する高エネルギー FADH2およびNADH分子を産生 解糖で処理されるグルコースの各分子について、36のatpsのネットは、好気性呼吸によって作成することができます。
放出されたエネルギーは、あなたの体を構成する細胞やシステムに電力を供給するために使用されます。 過剰または未利用のエネルギーは、後の使用のために脂肪またはグリコーゲンとして保存される。 炭水化物代謝は口の中で始まり、そこで酵素唾液アミラーゼは複雑な糖を単糖に分解し始める。 これらは腸の膜を渡って血流にそしてボディティッシュにそれから運ぶことができます。 細胞内では、六炭素糖であるグルコースが一連の反応によってより小さな糖に処理され、分子内に貯蔵されたエネルギーが放出される。 炭水化物異化の最初のステップは解糖であり、これはピルビン酸、NADHおよびATPを産生する。 嫌気性条件下では、ピルビン酸塩を乳酸塩に変換して解糖を維持することができる。 好気性条件下では、ピルビン酸はクエン酸サイクルまたはトリカルボン酸サイクルとも呼ばれるクレブスサイクルに入る。 ATPに加えて、Krebsサイクルは、より高エネルギーのATP分子を生成する酸化的リン酸化プロセスに電子を提供する高エネルギー FADH2およびNADH分子を産生 解糖で処理されるグルコースの各分子について、36のatpsのネットは、好気性呼吸によって作成することができます。
嫌気性条件下では、ATP産生は解糖によって生成されるものに限定される。 合計4つのAtpが解糖によって産生されるが、解糖を開始するには2つが必要であるため、2つのATP分子の正味収率がある。空腹時、飢餓、または低炭水化物食のような低グルコースの条件では、グルコースは乳酸塩、ピルビン酸塩、グリセロール、アラニン、またはグルタミン酸塩から合成 糖新生と呼ばれるこのプロセスは、解糖のほぼ逆であり、グルコースレベルが正常以下に低下すると、脳などのグルコース依存性器官のためのグルコース分子を作成するのに役立つ。
セルフチェック
前のセクションで説明したトピックをどれだけ理解しているかを確認するには、以下の質問に答えてください。
思考の質問
- グルコースがATPを生成するために代謝される方法を説明します。
- 細胞が血流からのグルコースの継続的な取り込みを確実にするために濃度勾配を作成するために採用するメカニズムを議論します。
用語集
多糖類:多くの単糖類で構成される複雑な炭水化物
単糖類:最小の単量体糖分子
唾液アミラーゼ: 唾液中に見出され、口の中で炭水化物の消化を開始する消化酵素
細胞呼吸:解糖、クレブスサイクル、および酸化的リン酸化を介してグルコース酸化からATPの生産
解糖:グルコースをピルビン酸に分解し、ATPを生成する一連の代謝反応
ピルビン酸:解糖の三炭素最終生成物とアセチルCoAに変換される出発物質
クレブスサイクルに入る: また、クエン酸サイクルまたはトリカルボン酸サイクルと呼ばれる、co2と高エネルギー FADH2、NADH、およびATP分子にピルビン酸を変換します
クエン酸サイ
グルコース-6-リン酸:解糖の最初のステップで生成されたリン酸化グルコース
ヘキソキナーゼ: 細胞酵素,ほとんどの組織で見つかった,細胞への取り込み時にグルコース-6-リン酸にグルコースを変換する
グルコキナーゼ:肝臓で見つかったcellularenzyme,細胞への取クレブス-サイクル: クエン酸サイクルまたはトリカルボン酸サイクルとも呼ばれ、ピルビン酸をCO2と高エネルギー FADH2、NADH、ATP分子に変換します
電子輸送鎖(ETC):電子が水を形成し、プロトン勾配を生成する一連の酸化還元反応を通過するATP産生経路
酸化的リン酸化:高エネルギー NADHとFADH2をATPに変換するプロセス
ATPシンターゼタンパク質:ATPを生成する細孔複合体
糖新生:ピルビン酸または他の分子からのグルコース合成のプロセス