Inleiding
succesvolle interactie met onze sensorische omgeving vereist een ingewikkelde balans van twee aandachtsselectiemechanismen—die van top-down en bottom-up. Op weg naar de producten gangpad van uw lokale supermarkt met het doel van het oppakken van enkele benodigde ingrediënten voor de mangosalade, betrekt inzet van vrijwillige doelgerichte, of top-down, aandacht systeem, zodat u actief zoeken naar alle benodigde ingrediënten onder de veelheid van producten keuzes. Echter, moet je een beltoon van een mobiele telefoon te horen, het zal waarschijnlijk uw aandacht vast te leggen en onderbreken uw zoekopdracht. Een dergelijke onderbreking vindt plaats op een bottom-up, of stimulus gedreven, manier waarbij een loutere ervaring van de stimulus, het feit dat de ring verschilt van andere geluiden in uw omgeving, acht het waardig van selectie. Het beschreven scenario onderstreept het belang van doelgerichte en stimulusgestuurde selectie voor gedrag, en wijst op een fijn evenwicht dat moet bestaan tussen de twee aandachtssystemen om “tunnelvisie” te voorkomen aan de ene kant en volledig onvermogen om zich op de andere te concentreren.
Top-Down en Bottom-Up selectie: Gedrag
verscheidene decennia van gedragsonderzoek zijn gewijd aan het aantonen dat de verdeling van aandacht kan worden gecontroleerd door intenties van de waarnemer en door de salience van de fysieke stimulus. Veel van gedragsmatig bewijs voor top-down en bottom-up attentionele allocatie is uitgebreid elders besproken (Johnston and Dark, 1986; Egeth and Yantis, 1997). Om samen te vatten, studies demonstreren effecten van top-down aandacht controle tonen aan dat de aandacht met succes kan worden toegewezen aan ruimtelijke locaties, functies, objecten, enz., na aanwezigheid van exogene of endogene cues (Eriksen en Hoffman, 1972; Posner, 1980; Posner et al.(Shaw, 1978; Moore en Egeth, 1998; Geng en Behrmann, 2002, 2005; Shomstein en Yantis, 2004a; Drummond en Shomstein, 2010). Bewijs dat bottom-up attentional allocation ondersteunt, is gebaseerd op verschillende attentional capture paradigma ‘ s, waarbij deelnemers betrokken zijn bij een top-down zoekopdracht en hun aandacht wordt omgeleid naar de taak-irrelevante stimuli, waaruit blijkt dat de aandacht wordt gevangen door feature singletons (unique item; Yantis and Jonides, 1990; Theeuwes, 1991; Folk et al., 2002) en abrupt onsets (Yantis en Jonides, 1984; Theeuwes, 1991; Koshino et al., 1992; Juola et al., 1995).
terwijl de meeste vroege studies zich concentreerden op het aantonen van bewijs voor top-down en bottom-up attentionele selectie, verleenden de meest recente studies hun focus op het onderzoeken van de interactie tussen de twee attentionele selectiesystemen. Deze lijn van onderzoek wordt gevoed door waarnemingen die om effectief te selecteren taak-relevante informatie (bijvoorbeeld, ingrediënten voor de salade) moet men actief remmen de taak-irrelevante informatie die anders de aandacht zou afleiden van de taak bij de hand. De keerzijde van deze logica is dat hoe minder Men is gericht op taakgerelateerde informatie, hoe meer capture zal volgen. Experimenteel is aangetoond dat de attentionele toestand van de waarnemer voorspelt wat voor soort informatie, en in welke mate, zal uiteindelijk de aandacht te vangen (Folk et al., 1992, 2002; Bacon and Egeth, 1994; Gibson and Kelsey, 1998). Bijvoorbeeld, Folk et al. (2002) toonde aan dat bij het zoeken naar een rode letter, een waarnemer gemakkelijker zal worden gevangen door een irrelevante stimulus in de periferie als die stimulus rood is, of overeenkomt met het doel sjabloon op een bepaalde manier. Aangezien de top-down control instellingen van de waarnemer zijn ingesteld om te zoeken naar een rode functie, elke stimulus die rood is waarschijnlijk de aandacht te vangen en mogelijk interfereren met top-down controle. Dus, met een capture-taak, kunnen aandachtszoekstrategieën van elkaar worden onderscheiden door de gelijkaardigheidsniveaus te variëren tussen de stimuluseigenschappen van het doel en de afleidingen. Hoe meer het doelwit lijkt op de distractor, hoe moeilijker het is voor de waarnemer om gevangenneming te vermijden.
de rol van de pariëtale kwab in Top-Down en Bottom-Up selectie: Neuroimaging
verschillende neuroimaging technieken leverden sterk bewijs voor de betrokkenheid van de pariëtale cortex in top-down en bottom-up oriëntering, met het bewijs uitgebreid elders besproken (Corbetta and Shulman, 2002, 2011; Behrmann et al., 2004). Er is aangetoond dat gebieden die het meest worden geactiveerd na top-down aanwijzingen om bij te wonen aan bepaalde locaties, functies, of objecten zich langs de dorsale delen van de pariëtale cortex bevinden. Dergelijke gebieden omvatten inferieure pariëtale lobule (IPL), dorsomediale regio ’s aangeduid als superior pariëtale lobule (SPL), evenals meer mediale regio’ s langs de precuneus gyrus (Yantis et al., 2002; Giesbrecht et al., 2003; Liu et al., 2003; Yantis and Serences, 2003; figuur 1). Verschillende top-down taken zijn shows om succesvol te betrekken dorsale gebieden van de pariëtale cortex, namelijk die waarbij ruimtelijke (Kastner et al., 1999; Corbetta et al., 2000; Hopfinger et al., 2000; Shomstein en Behrmann, 2006; Chiu en Yantis, 2009; Greenberg et al., 2010) en niet-ruimtelijke verschuivingen van aandacht (Giesbrecht et al., 2003; Yantis and Serences, 2003; Shomstein and Yantis, 2004b, 2006; Tamber-Rosenau et al., 2011).
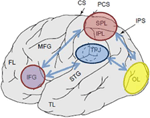
figuur 1. Schematische weergave van relevante anatomische oriëntatiepunten geprojecteerd op het laterale oppervlak van de menselijke hersenen. Superior pariëtale lobule (SPL) en inferior pariëtale lobule (IPL) zijn gebieden binnen het dorsale deel van de pariëtale cortex subserving top-down attentional orienting. Temporo-pariëtale kruising (TPJ) is een gebied binnen de ventrale pariëtale cortex subserving bottom-up attentional orienting. Beide, SPL en TPJ, worden verondersteld om controlesignalen die verantwoordelijk zijn voor de daaropvolgende aandacht modulaties waargenomen over sensorische gebieden, in dit geval modulerende (gelabeld met donkerblauwe pijlen) visueel opgeroepen activiteit in de occipitale kwab (OL). Bovendien worden gebieden langs de inferieure frontale gyrus (IFG) en inferieure frontale junctie (IFJ) verondersteld te dienen als convergentiegebieden voor stimulusgestuurde en top-down aandachtscontrole (gemarkeerd door lichtblauwe bi-directionele pijlen).
in een typische taak die gericht is op de attentionele toewijzing van bovenaf, worden individuen twee randgeplaatste RSVP-streams (rapid serial visual presentation) getoond en worden ze in eerste instantie geïnstrueerd om één stream te controleren op een cue (bijvoorbeeld een cijfer tussen de stroom letters). De identiteit van de cue geeft aan of het onderwerp aandacht moet houden op de huidige stroom of de aandacht moet verschuiven naar de andere stroom (Yantis et al., 2002; Yantis and Serences, 2003). In dergelijke paradigma ‘ s worden twee belangrijke bevindingen waargenomen. De eerste heeft te maken met verhoogde activering binnen de zintuiglijke gebieden die de op-het-moment bezochte plaats vertegenwoordigen (B.V., verhoogde activiteit binnen de linker primaire visuele gebieden wanneer de rechter RSVP-stroom wordt bijgewoond). Deze bevinding levert stevig bewijs dat deelnemers aanwezig zijn op een specifieke locatie en dat aandacht de kracht van de sensorische respons moduleert (zie Figuur 1; Moran and Desimone, 1985; O ‘ Raven et al., 1997). De tweede bevinding heeft te maken met de observatie dat dorsale gebieden van de pariëtale kwab selectief worden geactiveerd door verschuivingen van top-down aandacht. Er wordt opgemerkt dat de SPL / IPL-tijdloop van activiteit van voorbijgaande aard is, wat suggereert dat dit gebied van de pariëtale cortex de bron is van een kort aandachtscontrolesignaal om attente toestanden op een top-down manier te verschuiven (Yantis et al., 2002).
verschillende fMRI-studies hebben aangetoond dat bottom-up attention capture, gemedieerd door stimulus salience en/of relevantie, wordt ondersteund door de temporo-pariëtale kruising (TPJ; figuur 1). Bijvoorbeeld, wanneer proefpersonen een verandering in een visuele of auditieve stimulus, gelijktijdig gepresenteerd, bijwonen en controleren, wordt de activering van de TPJ-regio ‘ s van de pariëtale kwab versterkt. Naast de schijnbare gevoeligheid voor relevante stimuli, wordt TPJ ook geactiveerd als reactie op potentieel nieuwe (onverwachte of zeldzame) gebeurtenissen wanneer een organisme betrokken is in een neutrale gedragscontext of wanneer betrokken bij een taak (Marois et al., 2000; Downar et al., 2002; Serences et al., 2005; Corbetta et al., 2008; Asplund et al., 2010; Diquattro and Geng, 2011; Geng and Mangun, 2011). Deze activering vindt plaats onafhankelijk van de modaliteit (auditief, tactiel en visueel) waarin de input wordt geleverd, wat de multisensorische aard van TPJ weerspiegelt (maar zie Downar et al., 2001).
bij een typische taak die het neurale mechanisme van bottom-up attention capture onderzoekt, krijgen deelnemers een RSVP-stroom van items in het midden van het scherm te zien en wordt gevraagd om een vooraf gedefinieerd doel te identificeren (bijvoorbeeld rode letter te identificeren binnen een RSVP-stroom van witte niet-doelwitten). Een deel van de proeven bevat een taak-irrelevante saillent distractor gepresenteerd op verschillende tijdstippen voorafgaand aan het begin van het doel, terwijl andere proeven bevatten alleen de saillent distractor (dat wil zeggen, zonder het doel). “Target-distractor” proeven worden gebruikt om de omvang van de vangst te bepalen, waaruit blijkt dat de taak-irrelevante distractor in feite saillant is, waardoor een afname van de nauwkeurigheid van het doel wordt verkregen. De “distractor-in-isolation” – proeven worden gebruikt voor verdere analyses, aangezien dergelijke proeven het mogelijk maken om de activiteit te onderzoeken die wordt opgewekt door de meest opvallende distractor zonder contaminatie door de doelgerelateerde processen. Uit dergelijke paradigma ‘ s komen verschillende belangrijke bevindingen naar voren. Ten eerste, wanneer afleidingen ruimtelijk gescheiden zijn van de doellocatie, gaan afleidingen gepaard met verhoogde corticale activiteit in overeenkomstige gebieden van de sensorische cortex (bijvoorbeeld retinotopisch georganiseerde visuele cortex; zie Figuur 1). Dergelijke resultaten leveren sterk bewijs dat tijdens het vangen, ruimtelijke aandacht in feite wordt gevangen op de ruimtelijke locatie bezet door de distractor (Serences et al., 2005). Ten tweede, ventrale gebieden van de pariëtale cortex, voornamelijk binnen de TPJ worden selectief geactiveerd door bottom-up, onvrijwillige, verschuivingen van aandacht. Net zoals activiteit binnen de SPL voor de top-down oriëntering, is de tijdloop van activiteit waargenomen over TPJ van voorbijgaande aard, wat suggereert dat dit gebied de bron is van een kort aandachtscontrolesignaal om de aandacht op een bottom-up manier te verschuiven.
Opgemerkt dient te worden dat hoewel dit overzicht gericht is op het aanpakken van cognitieve functies van de posterieure pariëtale cortex, andere regio ‘ s, met name die binnen de frontale cortex, ook worden gerekruteerd voor top-down en bottom-up attentionele allocatie. Dergelijke gebieden omvatten de ventrale frontale cortex( VFC), de frontale oogvelden (FEF), inferieure frontale kruising (IFJ), en inferieure frontale gyrus (IFG; Corbetta and Shulman, 2002, 2011; Serences et al., 2005; Asplund et al., 2010; Diquattro and Geng, 2011).
de rol van de pariëtale kwab in Top-Down en Bottom-Up selectie: Neuropsychologie
Historisch baseerden onderzoekers zich kritisch op neuropsychologische studies van patiënten met hemispatiale verwaarlozing (een stoornis van ruimtelijke toewijzing van aandacht aan de linker hemi-ruimte) om inzicht te krijgen in cognitieve functies geassocieerd met de pariëtale kwab. In de klassieke neuropsychologische literatuur werd de pariëtale cortex, als geheel, algemeen beschouwd als de primaire laesieplaats voor hemispatiale verwaarlozing. Deze visie, in detail uitgewerkt door vroege onderzoekers (Critchley, 1953; McFire en Zangwill, 1960; Piercy, 1964) erkende duidelijk het verband tussen de pariëtale laesie en de daaruit voortvloeiende verwaarlozing. Dit perspectief werd grotendeels in de jaren 1980 gehouden toen Posner en collega ‘ s (1984) het verborgen visuospatiale cueing paradigma gebruikten om aan te tonen dat schade aan de pariëtale kwab een tekort in de “ontkoppeling” operatie veroorzaakt (het terugtrekken van de aandacht van de ene locatie en het verschuiven naar de andere) wanneer het doelwit contralateraal is met de laesie. Echter, ondanks deze grote vooruitgang in het begrijpen van de neurale basis van aandacht en in het bijzonder de “ontkoppeling” rol van de pariëtale cortex, hun bevindingen veronderstellen een enkele corticale site (pariëtale cortex) en een enkele functionele capaciteit (“ontkoppeling”). In tegenstelling tot deze meer monolithische benadering van de hersenen (pariëtale cortex) en gedrag (attentional disengagement), recente gedrag en neuroimaging werk (hierboven en elders besproken) suggereert dat zowel de corticale regio en de bijbehorende aandacht gedrag kan worden onderverdeeld in kwalitatief verschillende profielen.
gegeven de segregatie van de corticale netwerken in top-down en bottom-up processen, is een voor de hand liggende voorspelling dat schade aan superieure delen van de pariëtale lobule (subsuming SPL) een tekort in doelgerichte aandachtsoriëntatie zou moeten opleveren, terwijl schade aan de inferieure delen van de pariëtale lobule (subsuming TPJ) zou resulteren in een tekort geassocieerd met stimulus-gedreven aandacht vangen. In de mate dat deze hersengedrag correspondenties zijn onderzocht in de neuropsychologische literatuur, wordt deze voorspelling niet duidelijk bevestigd. Bijvoorbeeld, klinische symptomen van hemispatiale verwaarlozing zijn sterk geassocieerd met schade aan de inferieure delen van de pariëtale kwab, die TPJ omvat, in plaats van aan superieure delen zoals SPL (Friedrich et al., 1998; Shomstein et al., 2010; Corbetta and Shulman, 2011). Dit is enigszins in tegenspraak met de neuroimaging literatuur, die suggereert dat de rol van TPJ is in het vangen van de aandacht, in plaats van in de vrijwillige oriëntatie van de aandacht, het domein waarin verwaarlozing patiënten lijken te hebben de meeste moeite. Om de zaken nog ingewikkelder te maken, is opgemerkt dat letsels waarbij uitsluitend SPL betrokken is, slechts zelden klinisch bewijs van verwaarlozing opleveren (Vallar en Perani, 1986). Een andere recente studie met patiënten met laesies voornamelijk gecentreerd over TPJ en STG maar bewaard SPL, Corbetta et al. (2005) toonde aan dat ruimtelijke verwaarlozing, evenals het herstel ervan, werd geassocieerd met herstel van activiteit in zowel de ventrale temporo-pariëtale en dorsale pariëtale regio ‘ s (zie Corbetta en Shulman, 2011 voor een overzicht). Hoewel interessant en spannend in zijn conclusies, maakt deze laatste studie geen onderscheid tussen de relatieve bijdrage van dorsale en ventrale routes aan verschillende soorten aandacht, omdat patiënten alleen werden getest op een variant van de Posner covert spatial attention cuing task, taak die wordt verondersteld om zowel top-down als bottom-up aandacht oriëntering.
om onderscheid te maken tussen doelgestuurde aandachtscontrole en opvallende aandachtsopname en om hun mapping op respectievelijk de SPL en TPJ te onderzoeken, heeft recente studie twee gedragsparadigma ‘ s aangenomen, elk gericht op een van deze vormen van aandacht (Shomstein et al., 2010). Om de integriteit van top-down aandacht oriënterend in de patiënten te onderzoeken, werd een top-down taak gebruikt die deelnemers nodig heeft om ruimtelijke aandacht te verschuiven tussen de ruimtelijk gescheiden RSVP-streams (een taak die met succes is gebruikt om SPL-activering in fMRI-studies aan te tonen (Yantis et al., 2002)). Evenzo, om de bottom-up aandacht oriënterende capaciteiten van de patiënten te onderzoeken, een variant van Folk et al. (2002) contingent capture paradigma werd gebruikt in die deelnemers gedetecteerd doelen die verscheen bij fixatie terwijl taak-irrelevante kleur singletons werden geflitst in de periferie. De mate waarin taak-irrelevante afleidingen interfereren met de centrale detectietaak werd vervolgens gebruikt als een maat voor bottom-up attentional capture (Bacon and Egeth, 1994; Folk et al., 2002).
de voorspellingen waren als volgt: patiënten met laesies aan superieure delen van de pariëtale kwab (die SPL beïnvloeden) moeten worden aangetast bij de aandachtsoriëntatietaak van boven naar beneden (met behoud van prestaties bij de capture-taak) terwijl patiënten met laesies aan de inferieure delen van de pariëtale kwab (die TPJ beïnvloeden) moeten worden aangetast bij de capture-taak (met behoud van prestaties bij de top-down-taak). Een dubbele dissociatie van deze vorm getuigt niet alleen van de onafhankelijke componenten van aandacht, maar suggereert ook dat dergelijke attentionele componenten worden gemedieerd door onafhankelijke neurale mechanismen. Acht patiënten met visuo-ruimtelijke verwaarlozing werden gerekruteerd voor de studie en voltooiden twee taken, waarbij ofwel stimulusgestuurd ofwel doelgericht aandacht werd gericht. Op basis van hun gedragsprofiel werden patiënten in groepen gesorteerd en werd hun laesieoverlapping onderzocht (figuur 2A). Patiënten die moeilijkheden vertoonden met doelgerichte aandachtsoriëntatie, zoals gekwantificeerd door de top-down attentional index (figuur 2B), vertoonden laesieoverlapping gecentreerd over superieure delen van de pariëtale lobule (subsuming SPL) met spared inferior pariëtale lobule (TPJ). Patiënten met laesie overlap gecentreerd over de inferieure delen van de pariëtale lobule (subsuming TPJ) maar spared SPL uitgevoerd normaal op de doelgerichte oriënterende taak, terwijl de resterende immuun voor aandacht vangen (figuur 2C). De bevindingen van deze studie suggereren duidelijk dat SPL en TPJ anatomische gebieden zijn die noodzakelijkerwijs worden gerekruteerd met het oog op top-down en bottom-up oriëntering en dat schade aan SPL en TPJ leidt tot stoornissen van top-down en bottom-up oriëntering respectievelijk.
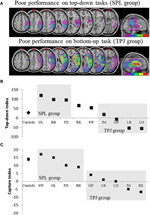
Figuur 2. Resultaten van de neuropsychologische studie gericht op het onderzoeken van de relatieve bijdrage van SPL en TPJ aan top-down en bottom-up oriëntering. (A) laesie overlappingen (paarse minimale overlapping; rode maximale overlapping) voor patiënten gegroepeerd op gedragstekorten in top-down aandacht oriëntering, gelabeld de SPL-groep (bovenpaneel); en patiënten gegroepeerd op gedragstekorten in bottom-up oriëntering, gelabeld de TPJ-groep (Onderpaneel). (B) Gedragsprestaties op de top-down taak samengevat met een “Top-down Index” die de verschillen kwantificeert tussen ruimtelijke top-down verschuivingen gemaakt van links naar rechts en vice versa. Controles en de TPJ lesioned-groep vertonen vergelijkbare efficiëntieverbeteringen bij het uitvoeren van ruimtelijke verschuivingen, terwijl patiënten met SPL-laesies een verminderde efficiëntie vertonen. Groep controle en individuele patiënt gegevens (gelabeld met patiënt initialen) zijn uitgezet op de abscis. (C)” Capture index ” is een maat voor bottom-up aandacht en kwantificeert de mate waarin taak-irrelevante afleidingen vangen aandacht weg van de taak. Besturingselementen en de SPL lesioned groep tonen vergelijkbare capture waarden, zodat beide groepen worden vastgelegd door de taak-irrelevante afleidingen. TPJ lesioned groep tonen veel gereduceerde capture index (niet te worden vastgelegd). Merk op dat patiënten werden geplaatst in de SPL of TPJ groep op basis van gedrag, in plaats van op basis van de laesie, dus let op de consistentie waarmee patiënten eindigen in de overeenkomstige groep.
interactie tussen Top-Down en Bottom-Up selectie
hoewel er blijkbaar een sterk verband bestaat tussen doelgerichte oriëntering en SPL en stimulusgestuurde oriëntering en TPJ, gegevens van Shomstein et al. (2010) patiëntstudie suggereert dat deze twee systemen niet volledig onafhankelijk zijn. Deze conclusie wordt ondersteund door de bevinding dat patiënten met SPL schade tentoongesteld een patroon van prestaties geëtiketteerd als “hyper capture.”In tegenstelling tot besturingselementen, voor wie alleen doel gekleurde distractor de aandacht trok (wat leidde tot een lagere doelnauwkeurigheid), bleken irrelevante gekleurde distractors ook af te leiden voor patiënten met SPL-laesie. Bovendien, terwijl voor controles aandacht werd gevangen door afleiding alleen wanneer ze voorafgaand aan het begin van het doel, voor patiënten met SPL laesies aandacht werd zelfs gevangen door afleiding gepresenteerd gelijktijdig met het doel. Dit prestatiepatroon kan worden verklaard door het volgende kader: SPL is verantwoordelijk voor top-down begeleiding van aandacht die het bepalen van de aspecten van de stimuli die taak relevant zijn omvat (bijvoorbeeld, zoeken naar rode doel; Corbetta en Shulman, 2002; Serences et al., 2005). Deze attentionele set beperkt dan TPJ, zodat de opname van aandacht mechanisme dat wordt gemedieerd door TPJ wordt alleen geactiveerd door de taak relevante informatie (bijvoorbeeld, rode afleidingen het vastleggen van de aandacht, en grijze afleidingen niet het vastleggen van de aandacht bij het zoeken naar een rode doel). De afwezigheid van SPL voorkomt de oprichting van een taak relevante aandacht set en dus elke stimulus, taak relevant of niet, wordt belangrijk geacht daarom het vangen van de aandacht (bijvoorbeeld, taak-irrelevante distractor vangen aandacht voor de SPL groep) willekeurig.
Er is gesuggereerd dat SPL en TPJ op minstens een van de twee mogelijke manieren kunnen samenwerken. De eerste mogelijkheid is dat TPJ dient als een waarschuwingssysteem dat gedragsmatig relevante stimuli detecteert maar de hoge ruimtelijke resolutie mist, dus wanneer een gedragsmatig relevante stimulus wordt gedetecteerd, wordt de precieze locatie geleverd door de SPL die ruimtelijke kaarten opslaat (Kastner et al., 1999; Wojciulik and Kanwisher, 1999; Bisley and Goldberg, 2003; Silver et al., 2005). Een verwante hypothetische mogelijkheid is dat het vangmechanisme (dat TPJ omvat) fungeert als een stroomonderbreker van voortdurende cognitieve activiteit wanneer een gedragsmatig relevante stimulus wordt gepresenteerd (Corbetta and Shulman, 2002, 2011). Het” hyper-capture ” patroon van activiteit waargenomen bij patiënten met bewaard gebleven TPJ maar lesioned SPL levert verder bewijs voor de hypothese dat TPJ ziet als het afgeven van een controle signaal dat de taak bij de hand beëindigt en dus dient als een stroomonderbreker (Corbetta and Shulman, 2002; Serences et al., 2005). Andere recente neuroimaging studies die gebruik maken van verschillende paradigma ‘ s hebben verder bewijs geleverd voor een interactieve relatie tussen de top-down en de bottom-up aandacht oriëntering, en vervolgens voor de relatie tussen SPL en TPJ (Serences et al., 2005; Asplund et al., 2010; Diquattro en Geng, 2011).
hoewel het bewijs voor een interactie tussen de twee attentionele systemen en de twee attentionele substraten (SPL en TPJ) sterk is, blijft onduidelijk of deze interactie direct is tussen SPL en TPJ of dat deze wordt bereikt via andere intermediaire regio ‘ s. Zoals eerder vermeld, richten top-down en bottom-up aandachtsoriëntatie netwerken zich op verschillende gebieden binnen de frontale cortex, dus is het redelijk om te veronderstellen dat de convergentie tussen de twee systemen kan worden bereikt via de frontale kwab. Twee recente studies die de interactie tussen top-down en bottom-up attentionele selectie onderzoeken, leverden bewijs voor de IFJ en IFG als mogelijke locaties van convergentie tussen stimulusgestuurde en doelgerichte selectie (Asplund et al., 2010; Diquattro and Geng, 2011). Het IFJ en het IFG lijken ideale kandidaten voor een dergelijke interactie gezien hun algemene betrokkenheid bij aandacht en cognitieve controle en hun betrokkenheid bij zowel ruimtelijke als niet-Ruimtelijke Selectie (Koechlin et al., 2003; Brass et al., 2005).
de rol van de pariëtale kwab in Top-Down en Bottom-Up selectie: Fysiologie
hoewel de nadruk in dit overzicht voornamelijk werd gelegd op studies bij de mens, is een grote schat aan kennis over de betrokkenheid van de pariëtale cortex bij de aandachtsoriëntatie afgeleid uit onderzoek naar de fysiologie van apen (zie recente review door Bisley and Goldberg, 2010). Echter, als het gaat om het onderzoeken van de relatieve bijdragen van verschillende regio ‘ s binnen de pariëtale cortex aan top-down en bottom-up aandacht Oriënteren, apenfysiologie literatuur schiet tekort. De belangrijkste reden hiervoor is dat er binnen de apenschors geen bewijs lijkt te zijn voor dezelfde segregatie van top-down en bottom-up controle. In plaats daarvan, laterale intrapariëtale gebied (LIP) oorspronkelijk dacht te zijn betrokken bij saccade planning (Gnadt en Andersen, 1988) is betrokken bij visuele aandacht en fungeert als een prioriteitskaart waarin externe stimuli worden weergegeven volgens hun gedragsprioriteit afgeleid op een top-down of bottom-up Manier (Colby en Goldberg, 1999; Bisley en Goldberg, 2003, 2010; Balan en Gottlieb, 2006; Ipata et al., 2006; Buschman and Miller, 2007; Gottlieb and Balan, 2010).
conclusie
hoewel veel minder bekend is over de menselijke pariëtale cortex dan die van de homologe Cortex van apen, zijn recente studies, waarbij gebruik wordt gemaakt van neuroimaging en neuropsychologische methoden, begonnen met het ophelderen van steeds fijnkorrelige functionele en structurele verschillen. Dit overzicht richtte zich op recente neuroimaging en neuropsychologische studies die de cognitieve rollen van dorsale en ventrale regio ‘ s van de pariëtale cortex in top-down en bottom-up attentionele oriëntering ophelderen, en op de interactie tussen de twee attentionele allocatiemechanismen.
belangenconflict verklaring
de auteur verklaart dat het onderzoek werd uitgevoerd zonder commerciële of financiële relaties die als een potentieel belangenconflict kunnen worden opgevat.
erkenningen
Dit werk werd ondersteund door de National Institutes of Health grant EY021644.
Asplund, C. L., Todd, J. J., Snyder, A. P., and Marois, R. (2010). Een centrale rol voor de laterale prefrontale cortex in doelgerichte en stimulusgestuurde aandacht. Nat. Neurowetenschappen. 13, 507–512.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Bacon, W. F., and Egeth, H. E. (1994). Overheersende stimulus-gedreven attention capture. Percept. Psychofys. 55, 485–496.
Pubmed Abstract / Pubmed Full Text
Balan, P. F., and Gottlieb, J. (2006). Integratie van exogene input in een dynamische salience map onthuld door het verstoren van de aandacht. J. Neurosci. 26, 9239–9249.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Behrmann, M., Geng, J. J., and Shomstein, S. (2004). Pariëtale cortex en aandacht. Curr. Opin. Neurobiol. 14, 212–217.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Bisley, J. W., and Goldberg, M. E. (2003). Neuronale activiteit in het laterale intrapariëtale gebied en ruimtelijke aandacht. Wetenschap 299, 81-86.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Bisley, J. W., and Goldberg, M. E. (2010). Aandacht, intentie en prioriteit in de pariëtale kwab. Annu. Rev. Neurosci. 33, 1–21.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Brass, M., Derrfuss, J., Forstmann, B., And von Cramon, D. Y. (2005). De rol van het inferieure frontale knooppuntgebied in cognitieve controle. Trends Cogn. Sci. 9, 314–316.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Buschman, T. J., and Miller, E. K. (2007). Top-down versus bottom-up controle van de aandacht in de prefrontale en posterieure pariëtale cortices. Wetenschap 315, 1860-1862.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Chiu, Y. C., and Yantis, S. (2009). Een domeinonafhankelijke bron van cognitieve controle voor taakverzamelingen: het verschuiven van ruimtelijke aandacht en het wisselen van categorisatieregels. J. Neurosci. 29, 3930–3938.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Colby, C. L., and Goldberg, M. E. (1999). Ruimte en aandacht in de pariëtale cortex. Annu. Rev. Neurosci. 22, 319–349.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., Kincade, J. M., Ollinger, J. M., McAvoy, M. P., and Shulman, G. L. (2000). Vrijwillige oriëntering wordt losgekoppeld van doeldetectie in de menselijke posterieure pariëtale cortex. Nat. Neurowetenschappen. 3, 292–297.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., Kincade, M. J., Lewis, C., Snyder, A. Z., and Sapir, A. (2005). Neurale basis en herstel van ruimtelijke aandachtstekorten in ruimtelijke verwaarlozing. Nat. Neurowetenschappen. 8, 1603–1610.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Corbetta, M., Patel, G., and Shulman, G. L. (2008). Het heroriënterende systeem van het menselijk brein: van omgeving naar theorie van de geest. Neuron 58, 306-324.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Corbetta, M., and Shulman, G. L. (2002). Controle van doelgerichte en stimulusgestuurde aandacht in de hersenen. Nat. Rev. Neurosci. 3, 201–215.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Corbetta, M., and Shulman, G. L. (2011). Ruimtelijke verwaarlozing en aandacht netwerken. Annu. Rev. Neurosci. 34, 569–599.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Critchley, M. (1953). De Pariëtale Kwabben. London, UK: Hafner Press.
Pubmed Abstract / Pubmed Full Text
Diquattro, N. E., and Geng, J. J. (2011). Contextuele kennis configureert aandacht control netwerken. J. Neurosci. 31, 18026–18035.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Downar, J., Crawley, A. P., Mikulis, D. J., and Davis, K. D. (2001). Het effect van taakrelevantie op de corticale respons op veranderingen in visuele en auditieve stimuli: een event-gerelateerde fMRI-studie. Neuroimage 14, 1256-1267.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Downar, J., Crawley, A. P., Mikulis, D. J., and Davis, K. D. (2002). Een corticaal netwerk gevoelig voor stimulus salience in een neutrale gedragscontext over meerdere zintuiglijke modaliteiten. J. Neurophysiol. 87, 615–620.
Pubmed Abstract / Pubmed Full Text
Drummond, L., and Shomstein, S. (2010). Object-gebaseerde aandacht: verschuivingen of onzekerheid? Attentie. Percept. Psychofys. 72, 1743–1755.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Egeth, H. E., and Yantis, S. (1997). Visuele aandacht: controle, representatie en tijdsverloop. Annu. Rev. Psychol. 48, 269–297.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Eriksen, C. W., and Hoffman, J. E. (1972). Temporele en ruimtelijke kenmerken van selectieve codering van visuele displays. Percept. Psychofys. 12, 201–204.
Folk, C. L., Leber, A. B., and Egeth, H. E. (2002). Ik liet je knipperen. Voorwaardelijke attention capture produceert een ruimtelijke knipoog. Percept. Psychofys. 64, 741–753.
Pubmed Abstract/Pubmed Full Text
Folk, C. L., Remington, R. W., and Johnston, J. C. (1992). Onvrijwillige covert oriëntatie is afhankelijk van aandacht controle instellingen. J. Exp. Psychol. Brom. Percept. Uitvoeren. 18, 1030–1044.
Pubmed Abstract / Pubmed Full Text
Friedrich, F. J., Egly, R., Rafal, R. D., and Beck, D. (1998). Spatial attention deficits in humans: a comparison of superior pariëtale and temporal-pariëtale junction laesies. Neuropsychologie 12, 193-207.
Pubmed Abstract / Pubmed Full Text
Geng, J. J., and Behrmann, M. (2002). Waarschijnlijkheid cuing van doellocatie vergemakkelijkt visueel zoeken impliciet bij normale deelnemers en patiënten met hemispatiale verwaarlozing. Psychol. sci. 13, 520–525.
Pubmed Abstract / Pubmed Full Text
Geng, J. J., and Behrmann, M. (2005). Ruimtelijke waarschijnlijkheid als een attentionele cue in visueel zoeken. Percept. Psychofys. 67, 1252–1268.
Pubmed Abstract / Pubmed Full Text
Geng, J. J., and Mangun, G. R. (2011). Rechts temporoparietal junction activering door een opvallende contextuele cue vergemakkelijkt doel discriminatie. Neuroimage 54, 594-601.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Gibson, B. S., and Kelsey, E. M. (1998). Stimulus-driven attentional capture is afhankelijk van attentional set voor displaywide visuele functies. J. Exp. Psychol. Brom. Percept. Uitvoeren. 24, 699–706.
Pubmed Abstract / Pubmed Full Text
Giesbrecht, B., Woldorff, M. G., Song, A. W., and Mangun, G. R. (2003). Neurale mechanismen van top-down controle tijdens ruimtelijke en functie aandacht. Neuroimage 19, 496-512.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Gnadt, J. W., and Andersen, R. A. (1988). Geheugen gerelateerde motorische planningsactiviteit in de achterste pariëtale cortex van makaak. Exp. Brain Res. 70, 216-220.
Pubmed Abstract / Pubmed Full Text
Gottlieb, J., and Balan, P. (2010). Aandacht als een beslissing in de informatieruimte. Trends Cogn. Sci. 14, 240–248.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Greenberg, A. S., Esterman, M., Wilson, D., Serences, J. T., and Yantis, S. (2010). Controle van ruimtelijke en functie-gebaseerde aandacht in frontoparietale cortex. J. Neurosci. 30, 14330–14339.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Hopfinger, J. B., Buonocore, M. H., and Mangun, G. R. (2000). De neurale mechanismen van top-down aandacht controle. Nat. Neurowetenschappen. 3, 284–291.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Ipata, A. E., Gee, A. L., Gottlieb, J., Bisley, J. W., and Goldberg, M. E. (2006). LIP reacties op een pop-out stimulus worden verminderd als het openlijk wordt genegeerd. Nat. Neurowetenschappen. 9, 1071–1076.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Johnston, W. A., and Dark, V. J. (1986). Selectieve Aandacht. Annu. Rev. Psychol. 37, 43–75.
Juola, J. F., Koshino, H., and Warner, C. B. (1995). Afwegingen tussen aandachtseffecten van ruimtelijke signalen en abrupte onsets. Percept. Psychofys. 57, 333–342.
Pubmed Abstract / Pubmed Full Text
Kastner, S., Pinsk, M. A., De Weerd, P., Desimone, R., and Ungereider, L. G. (1999). Verhoogde activiteit in de menselijke visuele cortex tijdens gerichte aandacht bij afwezigheid van visuele stimulatie. Neuron 22, 751-761.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Koechlin, E., Ody, C., and Kouneiher, F. (2003). De architectuur van cognitieve controle in de menselijke prefrontale cortex. Wetenschap 302, 1181-1185.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Koshino, H., Warner, C. B., and Juola, J. F. (1992). Relatieve effectiviteit van centrale, perifere en abrupte signalen in visuele aandacht. Q. J. Exp. Psychol. Een 45, 609-631.
Pubmed Abstract/Pubmed Full Text
Liu, T., Slotnick, S. D., Serences, J. T., and Yantis, S. (2003). Corticale mechanismen van functie-gebaseerde aandacht controle. Cereb. Cortex 13, 1334-1343.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Marois, R., Leung, H. C., and Gore, J. C. (2000). Een stimulusgestuurde benadering van objectidentiteit en locatieverwerking in het menselijk brein. Neuron 25, 717-728.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
McFire, J., and Zangwill, O. L. (1960). Visuo-constructieve beperkingen geassocieerd met laesies van de linker cerebrale hemisfeer. Brain 82, 243-260.
Moore, C. M., and Egeth, H. (1998). Hoe beïnvloedt feature-based attention visuele verwerking? J. Exp. Psychol. Brom. Percept. Uitvoeren. 24, 1296–1310.
Pubmed Abstract/Pubmed Full Text
Moran, J., and Desimone, R. (1985). Selectieve aandacht gates visuele verwerking in de extrastriate cortex. Wetenschap 229, 782-784.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
O ‘ Craven, K., Rosen, B. R., Kwong, K. K., Treisman, A., and Savoy, R. L. (1997). Vrijwillige aandacht moduleert fMRI-activiteit in menselijk MT-MST. Neuron 18, 591-598.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Piercy, M. (1964). The effects of cerebral laesies on intellectual function: een overzicht van de huidige onderzoekstrends. Br. J. Psychiatrie 110, 310-352.
Pubmed Abstract/Pubmed Full Text
Posner, M. I. (1980). Oriëntatie van de aandacht. Q. J. Exp. Psychol. 32, 3–25.
Pubmed Abstract/Pubmed Full Text
Posner, M. I., Snyder, C. R., and Davidson, B. J. (1980). Aandacht en detectie van signalen. J. Exp. Psychol. 109, 160–174.
Pubmed Abstract/Pubmed Full Text
Posner, M. I., Walker, J. A., Friedrich, F. J., and Rafal, R. D. (1984). Effecten van pariëtale schade op verborgen oriëntatie van de aandacht. J. Neurosci. 4, 1863–1874.
Pubmed Abstract/Pubmed Full Text
Serences, J. T., Shomstein, S., Leber, A. B., Golay, X., Egeth, H. E., and Yantis, S. (2005). Coördinatie van vrijwillige en stimulusgestuurde aandachtscontrole in de menselijke cortex. Psychol. Sci. 16, 114–122.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Shaw, M. L. (1978). Een capaciteitstoewijzingsmodel voor de reactietijd. J. Exp. Psychol. Brom. Percept. Uitvoeren. 4, 586–598.
Shomstein, S., en Behrmann, M. (2006). Corticale systemen die visuele aandacht bemiddelen voor zowel objecten als ruimtelijke locaties. Proc. Natl. Acad. Sci. U. S. A. 103, 11387-11392.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Shomstein, S., Lee, J., and Behrmann, M. (2010). Top-down en bottom-up attentionele begeleiding: onderzoek naar de rol van de dorsale en ventrale pariëtale cortices. Exp. Brain Res. 206, 197-208.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Shomstein, S., and Yantis, S. (2004a). Configurale en contextuele prioritering in object-gebaseerde aandacht. Psychon. Stier. Rev. 11, 247-253.
Pubmed Abstract/Pubmed Full Text
Shomstein, S., and Yantis, S. (2004b). Controle van aandacht verschuivingen tussen visie en auditie in de menselijke cortex. J. Neurosci. 24, 10702–10706.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Shomstein, S., and Yantis, S. (2006). Pariëtale cortex bemiddelt vrijwillige controle van ruimtelijke en niet-ruimtelijke auditieve aandacht. J. Neurosci. 26, 435–439.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Silver, M. A., Ress, D., and Heeger, D. J. (2005). Topografische kaarten van visuele ruimtelijke aandacht in de menselijke pariëtale cortex. J. Neurophysiol. 94, 1358–1371.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Tamber-Rosenau, B. J., Esterman, M., Chiu, Y.-C., and Yantis, S. (2011). Corticale mechanismen van cognitieve controle voor het verschuiven van aandacht in het gezichtsvermogen en werkgeheugen. J. Cogn. Neurowetenschappen. 23, 2905–2919.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Theeuwes, J. (1991). Exogene en endogene controle van de aandacht: het effect van visuele onsets en offsets. Percept. Psychofys. 49, 83–90.
Pubmed Abstract / Pubmed Full Text
Vallar, G., and Perani, D. (1986). De anatomie van unilaterale verwaarlozing na beroertes op de rechterhersenhelft. Een klinisch / CT-scan correlatieonderzoek bij de mens. Neuropsychologia 24, 609-622.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Wojciulik, E., and Kanwisher, N. (1999). De Algemene betrokkenheid van pariëtale bij visuele aandacht. Neuron 23, 747-764.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Yantis, S., and Jonides, J. (1984). Abrupte visuele onsets en selectieve aandacht: bewijs van visueel zoeken. J. Exp. Psychol. Brom. Percept. Uitvoeren. 10, 601–621.
Pubmed Abstract / Pubmed Full Text
Yantis, S., and Jonides, J. (1990). Abrupte visuele onsets en selectieve aandacht: vrijwillige versus automatische toewijzing. J. Exp. Psychol. Brom. Percept. Uitvoeren. 16, 121–134.
Pubmed Abstract / Pubmed Full Text
Yantis, S., Schwarzbach, J., Serences, J. T., Carlson, R. L., Steinmetz, M. A., Pekar, J. J., and Courtney, S. M. (2002). Voorbijgaande neurale activiteit in de menselijke pariëtale cortex tijdens ruimtelijke aandachtsverschuivingen. Nat. Neurowetenschappen. 5, 995–1002.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Yantis, S., and Serences, J. T. (2003). Corticale mechanismen van ruimtegebaseerde en objectgebaseerde aandachtscontrole. Curr. Opin. Neurobiol. 13, 187–193.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text