wprowadzenie
udana interakcja z naszym środowiskiem sensorycznym wymaga skomplikowanej równowagi dwóch uważnych mechanizmów selekcji—odgórnej i oddolnej. Kierując się do alejki produktów w lokalnym supermarkecie w celu zebrania kilku potrzebnych składników do sałatki z mango, angażuje się w wdrożenie dobrowolnego systemu ukierunkowanego na cel lub z góry na dół, uważnego, tak aby aktywnie szukać wszystkich wymaganych składników wśród wielu produktów do wyboru. Jeśli jednak usłyszysz dzwonek telefonu komórkowego, najprawdopodobniej przyciągnie twoją uwagę i przerwie wyszukiwanie. Takie przerwanie występuje w oddolnym lub napędzanym bodźcem sposobie, w którym samo znaczenie bodźca, fakt, że pierścień różni się od innych dźwięków w twoim otoczeniu, uznaje go za godny wyboru. Opisany scenariusz podkreśla znaczenie selekcji ukierunkowanej na cele i stymulowanej przez bodźce dla zachowania i wskazuje na delikatną równowagę, która musi istnieć między dwoma systemami uwagi, aby zapobiec „wizji tunelowej”z jednej strony i całkowitej niezdolności do skupienia się na drugiej.
wybór odgórny i odgórny: Zachowanie
kilkadziesiąt lat badań behawioralnych zostało poświęconych wykazaniu, że dystrybucja uwagi może być kontrolowana przez intencje obserwatora, jak również przez istotność bodźca fizycznego. Wiele dowodów behawioralnych dotyczących odgórnego i oddolnego przydzielania uwagi zostało szeroko przeanalizowanych w innych miejscach (Johnston and Dark, 1986; Egeth and Yantis, 1997). Podsumowując, badania wykazujące efekty odgórnej kontroli uwagi pokazują, że uwaga może być z powodzeniem przypisana do lokalizacji przestrzennych, cech, obiektów itp., po obecności egzogennych lub endogennych sygnałów (Eriksen i Hoffman, 1972; Posner, 1980; Posner i wsp., 1980), lub oczekiwania albo ustalone przez wcześniejszą wiedzę lub przez nieprzewidziane bodźce (Shaw, 1978; Moore and Egeth, 1998; Geng and Behrmann, 2002, 2005; Shomstein and Yantis, 2004a; Drummond and Shomstein, 2010). Dowody potwierdzające oddolną alokację uwagi opierały się na różnych paradygmatach przechwytywania uwagi, w których uczestnicy są zaangażowani w wyszukiwanie odgórne, a ich uwaga jest kierowana na bodźce nieistotne dla zadania, wykazując, że uwaga jest przechwytywana przez pojedyncze cechy (unique item; Yantis and Jonides, 1990; Theeuwes, 1991; Folk et al., 2002) i nagłe onsets (Yantis and Jonides, 1984; Theeuwes, 1991;Koshino et al., 1992; Juola et al., 1995).
podczas gdy większość wczesnych badań koncentrowała się na wykazaniu dowodów na odgórną i oddolną selekcję uważną, najnowsze badania skupiły się na badaniu, w jaki sposób oba systemy selekcji uważnej współdziałają ze sobą. Ta linia dochodzenia jest napędzany przez obserwacje, że w celu skutecznego wyboru istotnych dla zadania informacji (np. składniki sałatki) należy aktywnie hamować zadania nieistotne informacje, które w przeciwnym razie odwrócić uwagę od zadania pod ręką. Drugą stroną tej logiki jest to, że im mniej skupiamy się na informacjach związanych z zadaniami, tym więcej przechwytywania nastąpi. Wykazano eksperymentalnie, że stan uwagi obserwatora przewiduje, jaki rodzaj informacji i w jakim stopniu ostatecznie przyciągnie uwagę(Folk et al., 1992, 2002; Bacon and Egeth, 1994; Gibson and Kelsey, 1998). Na przykład Folk et al. (2002) pokazał, że podczas poszukiwania czerwonej litery, obserwator będzie łatwiej uchwycony przez nieistotny bodziec na peryferiach, jeśli bodziec ten jest czerwony lub pasuje do docelowego szablonu w jakiś sposób. Ponieważ odgórne ustawienia sterowania obserwatora są ustawione na wyszukiwanie czerwonej funkcji, każdy bodziec, który jest czerwony, prawdopodobnie przyciągnie uwagę i potencjalnie zakłóci kontrolę z góry na dół. Tak więc, dzięki zadaniu przechwytywania, strategie uważnego Wyszukiwania można odróżnić od siebie poprzez zmianę poziomów podobieństwa między właściwościami bodźców celu i rozpraszaczy. Im bardziej podobny jest cel do rozpraszacza, tym trudniej jest obserwatorowi uniknąć przechwycenia.
rola płata ciemieniowego w selekcji odgórnej i odgórnej: Neuroobrazowanie
różne techniki neuroobrazowania dostarczyły silnych dowodów na zaangażowanie kory ciemieniowej w orientację odgórną i oddolną, a dowody zostały szeroko przeanalizowane w innym miejscu (Corbetta and Shulman, 2002, 2011; Behrmann et al., 2004). Wykazano, że obszary najczęściej aktywowane po odgórnych wskazań, aby zająć się konkretnymi lokalizacjami, cechami lub przedmiotami, znajdują się wzdłuż grzbietowych części kory ciemieniowej. Takie obszary obejmują dolny płat ciemieniowy (IPL), regiony dorsomedialne określane jako górny płat ciemieniowy (SPL), a także bardziej przyśrodkowe regiony wzdłuż zakrętu precuneus (Yantis et al., 2002; Giesbrecht et al., 2003; Liu et al., 2003; Yantis and Serences, 2003; Rys. 1). Kilka odgórnych zadań zostało pokazanych, aby skutecznie zaangażować grzbietowe regiony kory ciemieniowej, a mianowicie te obejmujące przestrzenne (Kastner et al., 1999; Corbetta et al., 2000; Hopfinger et al., 2000; Shomstein and Behrmann, 2006; Chiu and Yantis, 2009; Greenberg et al., 2010), jak również nieprzestrzenne przesunięcia uwagi (Giesbrecht et al., 2003; Yantis and Serences, 2003; Shomstein and Yantis, 2004b, 2006; Tamber-Rosenau et al., 2011).
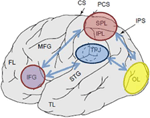
Rysunek 1. Schematyczne przedstawienie istotnych punktów anatomicznych rzutowanych na boczną powierzchnię ludzkiego mózgu. Górny płat ciemieniowy (SPL) i dolny płat ciemieniowy (IPL) są regionami w grzbietowej części kory ciemieniowej subserving z góry na dół Uwaga orientacja. Złącze skroniowo-ciemieniowe (TPJ) jest regionem w obrębie brzusznej kory ciemieniowej subserving bottom-up attentional orientation. Uważa się, że zarówno SPL, jak i TPJ wywołują sygnały kontrolne odpowiedzialne za późniejsze modulacje uwagi obserwowane w regionach zmysłowych, w tym przypadku modulujące (oznaczone ciemnoniebieskimi strzałkami) wizualnie wywołaną aktywność w płacie potylicznym (OL). Ponadto uważa się, że obszary wzdłuż gorszego zakrętu czołowego (IFG) i gorszego połączenia czołowego (IFJ) służą jako obszary konwergencji dla sterowanej bodźcem i kontroli z góry na dół (oznaczone jasnoniebieskimi dwukierunkowymi strzałkami).
w typowym zadaniu mającym na celu zaangażowanie odgórnego przydziału uwagi, jednostki są pokazywane dwa strumienie szybkiej szeregowej prezentacji wizualnej (RSVP) umieszczone peryferyjnie i są początkowo poinstruowane, aby monitorować jeden strumień dla cue (np. cyfra wśród strumienia liter). Tożsamość sygnalizacji wskazuje, czy podmiot musi zachować uwagę na bieżącym strumieniu, czy przenieść uwagę na inny strumień (Yantis et al., 2002; Yantis and Serences, 2003). W takich paradygmatach obserwuje się dwa główne odkrycia. Pierwszy ma do czynienia ze zwiększoną aktywacją w obszarach sensorycznych reprezentujących at-the-moment uczęszczanych lokalizacji (np, zwiększona aktywność w lewym podstawowych regionów wizualnych, gdy prawy strumień RSVP uczestniczy). To odkrycie dostarcza mocnych dowodów, że uczestnicy uczęszczają do określonej lokalizacji i że uwaga moduluje siłę odpowiedzi sensorycznej (patrz rysunek 1; Moran and Desimone, 1985; O ’ Craven et al., 1997). Drugie odkrycie ma związek z obserwacją, że grzbietowe obszary płata ciemieniowego są selektywnie aktywowane przez przesunięcia uwagi z góry na dół. Obserwuje się, że czasowy przebieg aktywności SPL/IPL ma charakter przejściowy, co sugeruje, że ten obszar kory ciemieniowej jest źródłem krótkiego uważnego sygnału sterującego do przesunięcia Stanów uważnych w sposób odgórny (Yantis et al., 2002).
kilka badań fMRI udokumentowało, że oddolne wychwytywanie uwagi, za pośrednictwem bodźca o znaczeniu i/lub istotności, jest podtrzymywane przez złącze skroniowo-ciemieniowe (TPJ; Fig.1). Na przykład, gdy badani zajmują się i monitorują zmianę bodźca wzrokowego lub słuchowego, prezentowaną jednocześnie, aktywacja regionów TPJ płata ciemieniowego jest wzmocniona. Oprócz pozornej wrażliwości na odpowiednie bodźce, TPJ jest również aktywowany w odpowiedzi na potencjalnie nowe (nieoczekiwane lub rzadkie) zdarzenia, gdy organizm jest zaangażowany w neutralnym kontekście behawioralnym lub gdy zaangażowany w zadanie (Marois et al., 2000; Downar et al., 2002; Serences et al., 2005; Corbetta et al., 2008; Asplund et al., 2010; Diquattro i Geng, 2011; Geng i Mangun, 2011). Aktywacja ta występuje niezależnie od modalności (słuchowej, dotykowej i wzrokowej), w której dane wejściowe są dostarczane, odzwierciedlając wieloczujnikowy charakter TPJ (ale patrz Downar et al., 2001).
w typowym zadaniu badającym neuronowy mechanizm oddolnego przechwytywania uwagi, uczestnicy są przedstawiani ze strumieniem RSVP przedmiotów w środku wyświetlacza i proszeni są o zidentyfikowanie wstępnie zdefiniowanego celu (np. zidentyfikuj czerwoną literę przedstawioną w strumieniu RSVP białych celów). Część badań zawiera nieistotny dla zadania element rozpraszający prezentowany w różnych odstępach czasu przed wystąpieniem celu, podczas gdy inne badania zawierają tylko element rozpraszający (tj. bez celu). Testy” Target-distractor ” są używane w celu określenia zakresu przechwytywania, pokazując, że nieistotny dla zadania rozpraszacz jest w rzeczywistości istotny, co prowadzi do zmniejszenia dokładności celu. Badania „rozpraszacz w izolacji” są wykorzystywane do dalszych analiz, ponieważ takie badania pozwalają na badanie aktywności wywoływanej do istotnego rozpraszacza bez zanieczyszczenia z procesów związanych z celem. Z takich paradygmatów wyłania się kilka ważnych wniosków. Po pierwsze, gdy rozpraszacze są przestrzennie oddzielone od docelowej lokalizacji, rozpraszaczom przechwytywania towarzyszy zwiększona aktywność korowa w odpowiednich regionach kory czuciowej (np. retinotopowo zorganizowana kora wzrokowa; patrz rysunek 1). Takie wyniki dostarczają mocnych dowodów na to, że podczas przechwytywania Uwaga przestrzenna jest w rzeczywistości przechwytywana do miejsca przestrzennego zajmowanego przez rozpraszacz (Serences et al., 2005). Po drugie, brzuszne regiony kory ciemieniowej, głównie w obrębie TPJ są selektywnie aktywowane przez oddolne, mimowolne, przesunięcia uwagi. Podobnie jak aktywność w SPL dla orientacji odgórnej, czasowy przebieg aktywności obserwowany nad TPJ ma charakter przejściowy, co sugeruje, że region ten jest źródłem krótkiego, uważnego sygnału sterującego, który przesuwa uwagę w sposób oddolny.
należy zauważyć, że podczas gdy ten przegląd koncentruje się na rozwiązywaniu funkcji poznawczych tylnej kory ciemieniowej, inne regiony, zwłaszcza te w korze czołowej, są również rekrutowane do uważnej alokacji odgórnej i oddolnej. Takie regiony obejmują przednią korę czołową (VFC), przednie pola oczu (FEF), inferior frontal junction (IFJ) i inferior frontal gyrus (IFG; Corbetta and Shulman, 2002, 2011; Serences et al., 2005; Asplund et al., 2010; Diquattro i Geng, 2011).
rola płata ciemieniowego w selekcji odgórnej i odgórnej: Neuropsychologia
historycy opierali się krytycznie na badaniach neuropsychologicznych pacjentów z zaniedbaniem hemispatial (zaburzeniem przestrzennego przydzielania uwagi do lewej pół-przestrzeni), aby uzyskać wgląd w funkcje poznawcze związane z płatem ciemieniowym. W klasycznej literaturze neuropsychologicznej, kora ciemieniowa, jako całość, była ogólnie uważana za główne miejsce uszkodzenia hemispatial zaniedbania. Pogląd ten, opracowany szczegółowo przez wczesnych badaczy (Critchley, 1953; McFire and Zangwill, 1960; Piercy, 1964) wyraźnie rozpoznał związek między uszkodzeniem ciemieniowym a wynikającym z tego zaniedbaniem. Ta perspektywa była w dużej mierze utrzymywana przez lata 80., kiedy Posner i współpracownicy (1984) wykorzystali tajny paradygmat wizualno-przestrzenny cueing, aby pokazać, że uszkodzenie płata ciemieniowego powoduje deficyt w operacji „odłączenia” (cofanie uwagi z jednego miejsca i przenoszenie jej w inne), gdy cel jest kontralateralny do zmiany. Jednak pomimo tego znacznego postępu w zrozumieniu neuronowej podstawy uwagi, a konkretnie roli „odłączenia” kory ciemieniowej, ich odkrycia zakładają pojedyncze miejsce korowe (kora ciemieniowa) i pojedynczą zdolność funkcjonalną („odłączenie”). W przeciwieństwie do tego bardziej monolitycznego podejścia do mózgu (kora ciemieniowa) i zachowania (uważne odłączenie), ostatnie prace behawioralne i neuroobrazowe (omówione powyżej i gdzie indziej) sugerują, że zarówno region korowy, jak i związane z nim zachowanie uważne mogą być podzielone na jakościowo różne profile.
biorąc pod uwagę segregację sieci korowych na procesy odgórne i oddolne, oczywistą prognozą jest, że uszkodzenie wyższych części płata ciemieniowego (subsuming SPL) powinno przynieść deficyt w ukierunkowanym na cel uważnym orientowaniu, podczas gdy uszkodzenie niższych części płata ciemieniowego (subsuming TPJ) spowodowałoby deficyt związany z wychwytywaniem uwagi przez bodźce. W zakresie, w jakim te korespondencje zachowania mózgu zostały zbadane w literaturze neuropsychologicznej, przewidywanie to nie jest oczywiście podtrzymywane. Na przykład, kliniczne objawy zaniedbania hemispatial są silnie związane z uszkodzeniem gorszych części płata ciemieniowego, który obejmuje TPJ, a nie do wyższych części, takich jak SPL (Friedrich et al., 1998; Shomstein et al., 2010; Corbetta i Shulman, 2011). Jest to nieco sprzeczne z literaturą neuroobrazowania, która sugeruje, że rola TPJ polega na przyciąganiu uwagi, a nie na dobrowolnym orientowaniu uwagi, domenie, w której pacjenci zaniedbujący wydają się mieć największe trudności. Aby jeszcze bardziej skomplikować sprawy, zauważono, że zmiany, które dotyczą wyłącznie SPL, rzadko dają kliniczne dowody zaniedbania (Vallar and Perani, 1986). Inne ostatnie badanie z udziałem pacjentów ze zmianami skoncentrowanymi głównie na TPJ i STG, ale zachowanymi SPL, Corbetta i wsp. (2005) wykazał, że zaniedbanie przestrzenne, jak również jego odzyskanie, było związane z przywróceniem aktywności zarówno w brzusznych regionach skroniowo-ciemieniowych, jak i grzbietowych (patrz Corbetta and Shulman, 2011 do przeglądu). Chociaż interesujące i ekscytujące w swoich wnioskach, to ostatnie badanie nie różnicuje względnego wkładu grzbietowej i brzusznej ścieżki do różnych rodzajów uwagi, ponieważ pacjenci byli testowani tylko na wariancie zadania Posner covert spatial attention cuing, zadania, które uważa się za angażujące zarówno odgórne, jak i oddolne orientowanie uwagi.
aby rozróżnić kontrolę celową i istotne wychwytywanie uwagi oraz zbadać ich mapowanie na odpowiednio SPL i TPJ, ostatnie badania przyjęły dwa paradygmaty behawioralne, z których każdy ukierunkowany jest na jedną z tych form uwagi (Shomstein et al., 2010). Aby zbadać integralność odgórnego orientowania uwagi u pacjentów, zastosowano zadanie odgórne wymagające od uczestników przesunięcia uwagi przestrzennej między oddzielonymi przestrzennie strumieniami RSVP (zadanie, które zostało z powodzeniem wykorzystane do wykazania aktywacji SPL w badaniach fMRI (Yantis et al., 2002)). Podobnie, w celu zbadania oddolnej uwagi zdolności orientacyjnych pacjentów, wariant Folk et al. (2002) paradygmat przechwytywania warunkowego został zastosowany, w którym uczestnicy wykrywali cele, które pojawiły się w fiksacji, podczas gdy nieistotne dla zadania kolorowe singletony były błyskane na peryferiach. Stopień, w jakim rozpraszacze nieistotne dla zadań zakłócają zadanie centralnego wykrywania, został następnie użyty jako miara oddolnego przechwytywania uwagi (Bacon and Egeth, 1994; Folk et al., 2002).
przewidywania były następujące: pacjenci ze zmianami w wyższych częściach płata ciemieniowego (wpływającymi na SPL) powinni być upośledzeni w zadaniu wychwytywania z góry na dół (przy zachowaniu wydajności w zadaniu wychwytywania), podczas gdy pacjenci ze zmianami w niższych częściach płata ciemieniowego (wpływającymi na TPJ) powinni być upośledzeni w zadaniu wychwytywania (przy zachowaniu wydajności w zadaniu wychwytywania z góry). Podwójna dysocjacja tej formy nie tylko świadczy o niezależnych składowych uwagi, ale również sugeruje, że takie składowe uwagi są pośredniczone przez niezależne mechanizmy neuronowe. Ośmiu pacjentów z zaniedbaniem wizualno-przestrzennym zostało zwerbowanych do badania i wykonało dwa zadania, wykorzystując orientację ukierunkowaną na bodźce lub cel. Na podstawie profilu behawioralnego pacjentów posortowano na grupy i zbadano pokrywanie się zmian chorobowych (rycina 2A). Pacjenci, którzy wykazywali trudności z ukierunkowanym na cel orientacją uważną, zgodnie z kwantyfikowanym wskaźnikiem uważności z góry na dół (Fig. 2b), przedstawiani byli z pokrywaniem się zmian wyśrodkowanych nad wyższymi częściami płata ciemieniowego (subsuming SPL) z oszczędzonym płatem ciemieniowym dolnym (TPJ). Pacjenci z uszkodzeniem pokrywają się wyśrodkowo nad dolnymi częściami płata ciemieniowego (subsuming TPJ), ale oszczędzają SPL wykonywaną normalnie w zadaniu zorientowania ukierunkowanego na cel, pozostając odpornymi na uważne wychwytywanie (Fig.2C). Wyniki tego badania wyraźnie sugerują, że SPL i TPJ są regionami anatomicznymi, które są koniecznie rekrutowane do celów orientacji odgórnej i oddolnej oraz że uszkodzenie SPL i TPJ prowadzi do zaburzeń orientacji odgórnej i oddolnej.
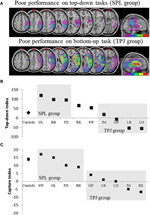
Rysunek 2. Wyniki badania neuropsychologicznego mającego na celu zbadanie względnego wpływu SPL i TPJ na Orientację odgórną i oddolną. (A) nakładanie się zmian (fioletowy Minimalne nakładanie się; czerwony maksymalne nakładanie się) dla pacjentów zgrupowanych według deficytów behawioralnych w orientacji uważnej z góry na dół, oznaczonych grupą SPL (górny panel); i pacjentów zgrupowanych według deficytów behawioralnych w orientacji oddolnej, oznaczonych grupą TPJ (dolny panel). (B) wydajność behawioralna zadania odgórnego podsumowana „indeksem odgórnym”, który określa ilościowo różnice między przestrzennymi przesunięciami odgórnymi od lewej do prawej i odwrotnie. Grupa kontrolna i grupa lesioned TPJ wykazują podobną wydajność w wykonywaniu przesunięć przestrzennych, podczas gdy pacjenci z uszkodzeniami SPL wykazują zmniejszoną wydajność. Grupa kontrolna i indywidualne dane pacjenta (oznaczone Inicjałami pacjenta) są wykreślone na abscissa. C) „Wskaźnik przechwytywania” jest miarą oddolnej uwagi i określa ilościowo stopień, w jakim rozpraszacze nieistotne dla zadania przyciągają uwagę z dala od zadania. Elementy sterujące i grupa SPL wykazują podobne wartości przechwytywania, tak że obie grupy są przechwytywane przez nieistotne dla zadania elementy rozpraszające. Grupa TPJ lesioned wykazuje znacznie zmniejszony indeks przechwytywania (Brak możliwości przechwycenia). Zauważ, że pacjenci zostali umieszczeni w grupie SPL lub TPJ na podstawie zachowania, a nie na podstawie zmiany, a zatem zwróć uwagę na spójność, z jaką pacjenci trafiają do odpowiedniej grupy.
interakcja między wyborem odgórnym i oddolnym
chociaż najwyraźniej istnieje silny związek między orientacją ukierunkowaną na cel i SPL oraz orientacją opartą na bodźcach i TPJ, dane z Shomstein et al. (2010) badanie pacjentów sugeruje, że te dwa systemy nie są całkowicie niezależne. Wniosek ten jest poparty stwierdzeniem, że pacjenci z uszkodzeniem SPL wykazywali wzorzec działania oznaczony jako ” Hyper capture.”W przeciwieństwie do kontrolerów, dla których tylko kolorowe rozpraszacze przyciągnęły uwagę (co prowadzi do mniejszej dokładności celu), nieistotne kolorowe rozpraszacze okazały się również rozpraszać pacjentów z uszkodzeniem SPL. Ponadto, podczas gdy w przypadku kontroli uwaga była przechwytywana przez rozpraszacze tylko wtedy, gdy poprzedzały one pojawienie się celu, w przypadku pacjentów ze zmianami SPL uwaga była nawet przechwytywana przez rozpraszacze prezentowane jednocześnie z celem. Ten wzorzec wydajności można wyjaśnić następującymi zasadami: SPL jest odpowiedzialny za odgórne kierowanie uwagą, która obejmuje określenie aspektów bodźców, które są istotne dla zadania (np search for red target; Corbetta and Shulman, 2002; Serences et al., 2005). Ten zestaw uwag ogranicza wtedy TPJ, tak że mechanizm przechwytywania uwagi, który jest pośredniczony przez TPJ, jest wyzwalany tylko przez informacje istotne dla zadania (np. czerwone rozpraszacze przechwytują uwagę, a szare rozpraszacze nie przechwytują uwagi podczas wyszukiwania czerwonego celu). Brak SPL uniemożliwia ustanowienie odpowiedniego zestawu uwag, a zatem każdy bodziec, zadanie istotne lub nie, jest uważany za ważny, dlatego przechwytywanie uwagi (np. rozpraszacz nieistotne zadanie przechwytywanie uwagi dla grupy SPL) bezkrytycznie.
zasugerowano, że SPL i TPJ mogą współdziałać na co najmniej jeden z dwóch możliwych sposobów. Pierwszą możliwością jest to, że TPJ służy jako system alarmowy, który wykrywa bodźce behawioralnie istotne, ale nie ma wysokiej rozdzielczości przestrzennej, dlatego gdy wykryty zostanie bodziec behawioralnie istotny, jego dokładna lokalizacja jest dostarczana przez SPL, który przechowuje mapy przestrzenne(Kastner et al., 1999; Wojciulik and Kanwisher, 1999; Bisley and Goldberg, 2003; Silver et al., 2005). Powiązana hipotetyczna możliwość polega na tym, że mechanizm przechwytywania (obejmujący TPJ) działa jako wyłącznik bieżącej aktywności poznawczej, gdy przedstawiony jest behawioralnie istotny bodziec (Corbetta and Shulman, 2002, 2011). Wzorzec aktywności” Hyper-capture ” obserwowany u pacjentów z zachowanym TPJ, ale uszkodzonym SPL dostarcza dalszych dowodów na hipotezę, że postrzega TPJ jako emitujący sygnał sterujący, który kończy zadanie, służąc w ten sposób jako wyłącznik (Corbetta and Shulman, 2002; Serences et al., 2005). Inne ostatnie badania neuroobrazowania wykorzystujące różne paradygmaty dostarczyły dalszych dowodów na interaktywną zależność między odgórnym i oddolnym orientowaniem uwagi, a następnie na związek między SPL i TPJ (Serences et al., 2005; Asplund et al., 2010; Diquattro i Geng, 2011).
chociaż dowody na interakcję między dwoma systemami uwagi i dwoma substratami uwagi (SPL i TPJ) są mocne, pozostaje niejasne, czy interakcja ta jest bezpośrednia między SPL i TPJ, czy odbywa się za pośrednictwem innych regionów pośrednich. Jak wspomniano wcześniej, odgórne i oddolne sieci uwagi orientacyjne angażują różne regiony w korze czołowej, dlatego uzasadnione jest hipoteza, że konwergencja między tymi dwoma systemami może być dokonana za pośrednictwem płata czołowego. Dwa ostatnie badania badające interakcję między odgórną i oddolną selekcją uważną dostarczyły dowodów na IFJ i IFG jako możliwe miejsca konwergencji między selekcją ukierunkowaną na bodźce i cel (Asplund et al., 2010; Diquattro i Geng, 2011). IFJ i IFG wydają się być idealnymi kandydatami do takiej interakcji, biorąc pod uwagę ich ogólne zaangażowanie w uwagę i kontrolę poznawczą, a także jego zaangażowanie w selekcji przestrzennej i nieprzestrzennej (Koechlin et al., 2003; Brass et al., 2005).
rola płata ciemieniowego w selekcji odgórnej i odgórnej: Fizjologia
podczas gdy nacisk w tej recenzji został położony głównie na badania na ludziach, ogromne bogactwo wiedzy na temat zaangażowania kory ciemieniowej w uważne orientowanie zostało zebrane z badań fizjologicznych małp (Zobacz najnowszą recenzję Bisley and Goldberg, 2010). Jednakże, jeśli chodzi o badanie względnego wkładu różnych regionów w korze ciemieniowej do uważnego orientowania odgórnego i oddolnego, Literatura fizjologii małp jest krótka. Głównym powodem tego jest to, że w korze małpiej nie wydaje się być dowodów na tę samą segregację kontroli odgórnej i oddolnej. Zamiast tego, boczny obszar wewnątrzparietalny (LIP) pierwotnie uważany za zaangażowany w planowanie saccade (Gnadt and Andersen, 1988) jest zaangażowany w uwagę wzrokową i działa jako mapa priorytetowa, w której bodźce zewnętrzne są reprezentowane zgodnie z ich priorytetem behawioralnym uzyskanym w sposób odgórny lub oddolny (Colby and Goldberg, 1999; Bisley and Goldberg, 2003, 2010; Balan and Gottlieb, 2006; Ipata et al., 2006; Buschman i Miller, 2007; Gottlieb i Balan, 2010).
wnioski
chociaż o ludzkiej korze ciemieniowej wiadomo znacznie mniej niż o homologicznej korze małpiej, ostatnie badania, wykorzystujące metody neuroobrazowania i neuropsychologiczne, zaczęły wyjaśniać coraz bardziej drobnoziarniste różnice funkcjonalne i strukturalne. Przegląd ten koncentrował się na najnowszych badaniach neuroobrazowania i neuropsychologicznych wyjaśniających poznawcze role grzbietowych i brzusznych regionów kory ciemieniowej w orientacji uważnej z góry i z dołu do góry oraz na interakcji między dwoma mechanizmami alokacji uwagi.
Oświadczenie o konflikcie interesów
autor oświadcza, że badanie zostało przeprowadzone przy braku jakichkolwiek relacji handlowych lub finansowych, które mogłyby być interpretowane jako potencjalny konflikt interesów.
podziękowania
praca ta została wsparta przez Grant Narodowego Instytutu Zdrowia EY021644.
Asplund, C. L., Todd, J. J., Snyder, A. P., and Marois, R. (2010). Centralna rola bocznej kory przedczołowej w ukierunkowanej na cel i stymulowanej uwadze. Nat. Neurosci. 13, 507–512.
PubMed Abstract | PubMed Full Text | CrossRef Full Text
Bacon, W. F., And Egeth, H. E. (1994). Nadrzędne wychwytywanie uwagi przez bodźce. Percept. Psychofy. 55, 485–496.
PubMed Streszczenie | PubMed Pełny tekst
Balan, P. F., and Gottlieb, J. (2006). Integracja egzogennego wejścia w dynamiczną mapę widoczności ujawnioną przez niepokojącą uwagę. J. Neurosci. 26, 9239–9249.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Behrmann, M., Geng, J. J., and Shomstein, S. (2004). Kora ciemieniowa i uwaga. Curr. Opin. Neurobiol. 14, 212–217.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Bisley, J. W., and Goldberg, M. E. (2003). Aktywność neuronalna w bocznym obszarze wewnątrzgałkowym i uwaga przestrzenna. Nauka 299, 81-86.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Bisley, J. W., and Goldberg, M. E. (2010). Uwaga, intencja i priorytet w płacie ciemieniowym. Annu. Ks. Neurosci. 33, 1–21.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Brass, M., Derrfuss, J., Forstmann, B., and von Cramon, D. Y. (2005). Rola dolnego obszaru złącza czołowego w kontroli poznawczej. Trendy Cogn. Sci. 9, 314–316.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Buschman, T. J., and Miller, E. K. (2007). Top-down kontra bottom-up kontrola uwagi w przedczołowej i tylnej kory ciemieniowej. Nauka 315, 1860-1862.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Chiu, Y. C., and Yantis, S. (2009). Niezależne od domeny źródło kontroli poznawczej dla zestawów zadań: przesunięcie uwagi przestrzennej i zmiana reguł kategoryzacji. J. Neurosci. 29, 3930–3938.
PubMed Abstract | PubMed Full Text | CrossRef Full Text
Przestrzeń i uwaga w korze ciemieniowej. Annu. Ks. Neurosci. 22, 319–349.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Corbetta, M., Kincade, J. M., Ollinger, J. M., McAvoy, M. P., and Shulman, G. L. (2000). Dobrowolne orientowanie jest oddzielone od wykrywania celu w ludzkiej tylnej korze ciemieniowej. Nat. Neurosci. 3, 292–297.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Corbetta, M., Kincade, M. J., Lewis, C., Snyder, A. Z., and Sapir, A. (2005). Podstawy neuronowe i odzyskiwanie przestrzennych deficytów uwagi w zaniedbaniu przestrzennym. Nat. Neurosci. 8, 1603–1610.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Corbetta, M., Patel, G., and Shulman, G. L. (2008). System reorientacji ludzkiego mózgu: od środowiska do teorii umysłu. Neuron 58, 306-324.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Corbetta, M., and Shulman, G. L. (2002). Kontrola ukierunkowanej na cel i stymulowanej uwagi w mózgu. Nat. Ks. Neurosci. 3, 201–215.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Corbetta, M., and Shulman, G. L. (2011). Przestrzenne zaniedbania i sieci uwagi. Annu. Ks. Neurosci. 34, 569–599.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Critchley, M. (1953). Płaty Ciemieniowe. Londyn, Wielka Brytania: Hafner Press.
PubMed Streszczenie | PubMed Pełny tekst
Diquattro, N. E., and Geng, J. J. (2011). Wiedza kontekstowa konfiguruje sieci kontroli uwagi. J. Neurosci. 31, 18026–18035.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Downar, J., Crawley, A. P., Mikulis, D. J., and Davis, K. D. (2001). The effect of task relevance on the cortical response to changes in visual and auditory stimuli: an event-related fMRI study. Neuroimage 14, 1256-1267.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Downar, J., Crawley, A. P., Mikulis, D. J., and Davis, K. D. (2002). Sieć korowa wrażliwa na znaczenie bodźców w neutralnym kontekście behawioralnym w wielu modalnościach sensorycznych. J. Neurofizjol. 87, 615–620.
PubMed Streszczenie | PubMed Pełny tekst
Uwaga oparta na obiektach: przesunięcie czy niepewność? Uwaga. Percept. Psychofy. 72, 1743–1755.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Egeth, H. E., and Yantis, S. (1997). Uwaga wzrokowa: Kontrola, reprezentacja i przebieg czasu. Annu. Ks. Psychol. 48, 269–297.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Eriksen, C. W., and Hoffman, J. E. (1972). Czasowa i przestrzenna charakterystyka selektywnego kodowania z wyświetlaczy wizualnych. Percept. Psychofy. 12, 201–204.
Folk, C. L., Leber, A. B., and Egeth, H. E. (2002). Mrugnąłeś! Warunkowe Przechwytywanie z uwagą powoduje mrugnięcie przestrzenne. Percept. Psychofy. 64, 741–753.
PubMed Streszczenie | PubMed Pełny tekst
Folk, C. L., Remington, R. W., and Johnston, J. C. (1992). Mimowolne tajne orientowanie zależy od uważnych ustawień kontroli. J. Exp. Psychol. Hum. Percept. Wykonaj. 18, 1030–1044.
PubMed Streszczenie | PubMed Pełny tekst
Friedrich, F. J., Egly, R., Rafal, R. D., and Beck, D. (1998). Przestrzenne deficyty uwagi u ludzi: porównanie wyższych zmian ciemieniowych i skroniowo-ciemieniowych. Neuropsychologia 12, 193-207.
PubMed Streszczenie | PubMed Pełny tekst
Geng, J. J., and Behrmann, M. (2002). Prawdopodobieństwo cuing docelowej lokalizacji ułatwia wyszukiwanie wzrokowe pośrednio u normalnych uczestników i pacjentów z zaniedbaniem hemispatial. Psychol. sci. 13, 520–525.
PubMed Streszczenie | PubMed Pełny tekst
Geng, J. J., and Behrmann, M. (2005). Prawdopodobieństwo przestrzenne jako uwaga w poszukiwaniu wizualnym. Percept. Psychofy. 67, 1252–1268.
PubMed Streszczenie | PubMed Pełny tekst
Geng, J. J., and Mangun, G. R. (2011). Aktywacja prawego połączenia skroniowo-cieczowego za pomocą znaczącego sygnału kontekstowego ułatwia dyskryminację celu. Neuroimage 54, 594-601.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Gibson, B. S., and Kelsey, E. M. (1998). Bodziec-driven attentional capture jest uzależnione od attentional zestaw dla funkcji wizualnych displaywide. J. Exp. Psychol. Hum. Percept. Wykonaj. 24, 699–706.
PubMed Streszczenie | PubMed Pełny tekst
Giesbrecht, B., Woldorff, M. G., Song, A. W., and Mangun, G. R. (2003). Neuronowe mechanizmy kontroli odgórnej podczas uwagi przestrzennej i Cechowej. Neuroimage 19, 496-512.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Gnadt, J. W., and Andersen, R. A. (1988). Aktywność planowania motorycznego związana z pamięcią w tylnej korze ciemieniowej makaka. Exp. Brain Res. 70, 216-220.
PubMed Streszczenie | PubMed Pełny tekst
Gottlieb, J., and Balan, P. (2010). Uwaga jako decyzja w przestrzeni informacyjnej. Trendy Cogn. Sci. 14, 240–248.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Greenberg, A. S., Esterman, M., Wilson, D., Serences, J. T., and Yantis, S. (2010). Kontrola uwagi przestrzennej i Cechowej w korze czołowo-cieczowej. J. Neurosci. 30, 14330–14339.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Hopfinger, J. B., Buonocore, M. H., and Mangun, G. R. (2000). Neuronowe mechanizmy kontroli z góry na dół. Nat. Neurosci. 3, 284–291.
PubMed Abstract | PubMed Full Text | CrossRef Full Text
, Gee, A. L., Gottlieb, J., Bisley, J. W., and Goldberg, M. E. (2006). Reakcje warg na bodziec popout są zmniejszone, jeśli jest on jawnie ignorowany. Nat. Neurosci. 9, 1071–1076.
PubMed Abstract | PubMed Full Text | CrossRef Full Text
Selektywna Uwaga. Annu. Ks. Psychol. 37, 43–75.
Juola, J. F., Koshino, H., and Warner, C. B. (1995). Kompromisy między uważnymi efektami sygnałów przestrzennych a nagłymi zestawami. Percept. Psychofy. 57, 333–342.
PubMed Streszczenie | PubMed Pełny tekst
Kastner, S., Pinsk, M. A., De Weerd, P., Desimone, R., and Ungerleider, L. G. (1999). Zwiększona aktywność ludzkiej kory wzrokowej podczas ukierunkowanej uwagi w przypadku braku stymulacji wzrokowej. Neuron 22, 751-761.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Koechlin, E., Ody, C., and Kouneiher, F. (2003). Architektura kontroli poznawczej w korze przedczołowej człowieka. Nauka 302, 1181-1185.
PubMed Abstract | PubMed Full Text | CrossRef Full Text
, Warner, C. B., and Juola, J. F. (1992). Względna skuteczność sygnałów ośrodkowego, obwodowego i nagłego początku w zwróceniu uwagi wzrokowej. Q. J. Exp. Psychol. A 45, 609-631.
PubMed Streszczenie | PubMed Pełny tekst
Liu, T., Slotnick, S. D., Serences, J. T., and Yantis, S. (2003). Korowe mechanizmy kontroli uwagi opartej na cechach. Mózg. Cortex 13, 1334-1343.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Marois, R., Leung, H. C., and Gore, J. C. (2000). Stymulowane podejście do przetwarzania tożsamości i lokalizacji obiektów w ludzkim mózgu. Neuron 25, 717-728.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
McFire, J., and zangwill, O. L. (1960). Wizualno-konstruktywne niepełnosprawności związane ze zmianami lewej półkuli mózgowej. Brain 82, 243-260.
Moore, C. M., and Egeth, H. (1998). W jaki sposób UWAGA oparta na funkcjach wpływa na przetwarzanie obrazu? J. Exp. Psychol. Hum. Percept. Wykonaj. 24, 1296–1310.
PubMed Streszczenie | PubMed Pełny tekst
, and Desimone, R. (1985). Selektywna Uwaga gates przetwarzania wizualnego w korze extrastriate. Nauka 229, 782-784.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
O ’ Craven, K., Rosen, B. R., Kwong, K. K., Treisman, A., and Savoy, R. L. (1997). Dobrowolna Uwaga moduluje aktywność fMRI u człowieka MT-MST. Neuron 18, 591-598.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Wpływ zmian mózgowych na funkcje intelektualne: przegląd aktualnych trendów badawczych. Br. J. Psychiatra 110, 310-352.
PubMed Streszczenie | PubMed Pełny tekst
Posner, M. I. (1980). Orientacja uwagi. Q. J. Exp. Psychol. 32, 3–25.
PubMed Streszczenie | PubMed Pełny tekst
Posner, M. I., Snyder, C. R., and Davidson, B. J. (1980). Uwaga i wykrywanie sygnałów. J. Exp. Psychol. 109, 160–174.
PubMed Streszczenie | PubMed Pełny tekst
Posner, M. I., Walker, J. A., Friedrich, F. J., and Rafal, R. D. (1984). Wpływ urazu ciemieniowego na Ukryte orientowanie uwagi. J. Neurosci. 4, 1863–1874.
PubMed Streszczenie | PubMed Pełny tekst
Serences, J. T., Shomstein, S., Leber, A. B., Golay, X., Egeth, H. E., and Yantis, S. (2005). Koordynacja dobrowolnej i stymulowanej uwagi kontroli w korze ludzkiej. Psychol. Sci. 16, 114–122.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Model alokacji zdolności na czas reakcji. J. Exp. Psychol. Hum. Percept. Wykonaj. 4, 586–598.
Shomstein, S., and Behrmann, M. (2006). Systemy korowe pośredniczące w wizualnej obserwacji zarówno obiektów, jak i miejsc przestrzennych. Proc. Natl. Acad. Sci. U. S. A. 103, 11387-11392.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Shomstein, S., Lee, J., and Behrmann, M. (2010). Top-down I bottom-up attentional guidance: badanie roli grzbietowej i brzusznej kory ciemieniowej. Exp. Brain Res. 206, 197-208.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Shomstein, S., and Yantis, S. (2004a). Konfigurowalne i kontekstowe priorytetyzowanie uwagi obiektowej. Psychon. Bzdura. Rev. 11, 247-253.
PubMed Streszczenie | PubMed Pełny tekst
Shomstein, S., and Yantis, S. (2004b). Kontrola przesunięć uwagi między widzeniem a słuchem w ludzkiej korze mózgowej. J. Neurosci. 24, 10702–10706.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Shomstein, S., and Yantis, S. (2006). Kora ciemieniowa pośredniczy w dobrowolnej kontroli przestrzennej i bezprzestrzennej uwagi słuchowej. J. Neurosci. 26, 435–439.
PubMed Abstract | PubMed Full Text | CrossRef Full Text
Silver, M. A., Ress, D., and Heeger, D. J. (2005). Mapy topograficzne wizualnej uwagi przestrzennej w korze ciemieniowej człowieka. J. Neurofizjol. 94, 1358–1371.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Tamber-Rosenau, B. J., Esterman, M., Chiu, Y.-C., and Yantis, S. (2011). Korowe mechanizmy kontroli poznawczej dla przesunięcia uwagi w widzeniu i pamięci roboczej. J. Cogn. Neurosci. 23, 2905–2919.
PubMed Abstract | PubMed Full Text | CrossRef Full Text
Theeuwes, J. (1991). Egzogenna i endogenna Kontrola uwagi: efekt wizualnych onsets i offsets. Percept. Psychofy. 49, 83–90.
PubMed Streszczenie / PubMed Pełny tekst
Vallar, G., and Perani, D. (1986). Anatomia jednostronnego zaniedbania po zmianach udarowych prawej półkuli. Badanie korelacji klinicznej / tomografii komputerowej u ludzi. Neuropsychologia 24, 609-622.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Wojciulik, E., and Kanwisher, N. (1999). Ogólność zaangażowania ciemieniowego w uwagę wzrokową. Neuron 23, 747-764.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Yantis, S., and Jonides, J. (1984). Nagłe wizualne zdarzenia i selektywna Uwaga: dowody z wyszukiwania wizualnego. J. Exp. Psychol. Hum. Percept. Wykonaj. 10, 601–621.
PubMed Streszczenie/PubMed Pełny tekst
Yantis, S., and Jonides, J. (1990). Nagłe wizualne ustawienia i selektywna Uwaga: dobrowolne kontra automatyczne przydzielanie. J. Exp. Psychol. Hum. Percept. Wykonaj. 16, 121–134.
PubMed Streszczenie | PubMed Pełny tekst
Yantis, S., Schwarzbach, J., Serences, J. T., Carlson, R. L., Steinmetz, M. A., Pekar, J. J., and Courtney, S. M. (2002). Przemijająca aktywność nerwowa w ludzkiej korze ciemieniowej podczas przestrzennych przesunięć uwagi. Nat. Neurosci. 5, 995–1002.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst
Yantis, S., and Serences, J. T. (2003). Korowe mechanizmy kontroli uwagi opartej na przestrzeni i obiektowej. Curr. Opin. Neurobiol. 13, 187–193.
PubMed Streszczenie | PubMed Pełny tekst | CrossRef Pełny tekst