Introdução
bem-Sucedida interação com o ambiente sensorial requer um complexo equilíbrio de dois atencional mecanismos de seleção—o de cima para baixo e de baixo para cima. Se dirigir até a produzir o corredor de seu supermercado local com o objetivo de pegar alguns ingredientes necessários para a manga salada, envolve a implantação de voluntários meta-dirigida, ou de cima para baixo, atencional sistema, para que você procurar activamente todos os ingredientes necessários entre a multidão de produzir escolhas. No entanto, se ouvir o som de um telemóvel, provavelmente irá captar a sua atenção e interromper a sua busca. Tal interrupção ocorre em uma forma bottom-up, ou impulsionada pelo estímulo, em que uma mera experiência do estímulo, o fato de que o anel é diferente de outros sons em seu ambiente, o considera digno de seleção. O cenário descrito sublinha a importância da seleção de comportamento orientada para o objetivo e impulsionada pelo estímulo, e aponta para um equilíbrio fino que tem de existir entre os dois sistemas de atenção para evitar a “visão do túnel”, por um lado, e completa incapacidade de se concentrar no outro.selecção de
de cima para baixo e de baixo para cima: Comportamento
várias décadas de pesquisa comportamental têm sido dedicadas a demonstrar que a distribuição da atenção pode ser controlada por intenções do observador, bem como pela experiência do estímulo físico. Muitas das evidências comportamentais para alocação de atenção top-down e bottom-up foram revisadas extensivamente em outros lugares (Johnston e Dark, 1986; Egeth e Yantis, 1997). Para resumir, estudos que demonstram os efeitos do controlo de atenção de cima para baixo mostram que a atenção pode ser atribuída com sucesso a localizações espaciais, características, objectos, etc., na sequência da presença de sinais exógenos ou endógenos (Eriksen e Hoffman, 1972; Posner, 1980; Posner et al., 1980), ou expectativas definidos pelo conhecimento prévio ou por contingências de estímulo (Shaw, 1978; Moore e Egeth, 1998; Geng e Behrmann, 2002, 2005; Shomstein e Yantis, 2004a; Drummond e Shomstein, 2010). Provas de baixo para cima atencional de alocação foi invocado várias atencional de captura de paradigmas, em que os participantes estão envolvidos em um top-down de pesquisa e sua atenção é desviada para a tarefa-estímulos irrelevantes, demonstrando que a atenção é capturada pelo recurso de gestações únicas (unique item; Yantis e Jonides, 1990; Theeuwes, 1991; Folk et al., 2002) and abrupt onsets (Yantis and Jonides, 1984; Theeuwes, 1991; Koshino et al., 1992; Juola et al., 1995).enquanto a maioria dos estudos iniciais se concentrou na demonstração de evidências para a seleção de atenção top-down e bottom-up, os estudos mais recentes mudaram seu foco para examinar como os dois sistemas de seleção de atenção interagem. Esta linha de investigação é alimentada por observações que, a fim de selecionar eficazmente a informação relevante tarefa (por exemplo, ingredientes para a salada) deve-se ativamente inibir a tarefa-informação irrelevante que de outra forma desviaria a atenção da tarefa em mãos. O outro lado desta lógica, é que quanto menos um está focado em informações relacionadas com tarefas, mais Captura irá se seguir. Foi demonstrado experimentalmente que o estado de atenção do observador prevê que tipo de informação, e em que medida, acabará por atrair a atenção (Folk et al., 1992, 2002; Bacon and Egeth, 1994; Gibson and Kelsey, 1998). Por exemplo, Folk et al. (2002) mostrou que ao procurar por uma letra vermelha, um observador será mais facilmente capturado por um estímulo irrelevante na periferia se esse estímulo for vermelho, ou se corresponder de alguma forma ao modelo-alvo. Uma vez que as configurações de controle de cima para baixo do observador estão definidas para procurar por uma característica vermelha, qualquer estímulo que seja vermelho é provável que capture atenção e potencialmente interfira com o controle de cima para baixo. Assim, com uma tarefa de captura, as estratégias de busca de atenção podem ser distinguidas umas das outras, variando os níveis de similaridade entre as propriedades de estímulo do alvo e dos distractores. Quanto mais similar o alvo é ao distrator, mais difícil é para o observador evitar a captura.
o papel do Lobo Parietal na selecção de cima para baixo e de baixo para cima: Neuroimagem
Diversas técnicas de neuroimagem forte evidência para o envolvimento do córtex parietal no top-down e bottom-up, de orientação, com a evidência analisou amplamente em outros lugares (Corbetta e Shulman, 2002, 2011; Behrmann et al., 2004). Tem sido demonstrado que áreas mais comumente ativadas seguindo pistas top-down para atender a locais, características ou objetos particulares estão localizados ao longo das partes dorsais do córtex parietal. Tais áreas incluem o lobo parietal inferior( IPL), as regiões dorsomediais referidas como lobo parietal superior (SPL), bem como mais regiões mediais ao longo do giro precuneus (Yantis et al., 2002; Giesbrecht et al., 2003; Liu et al., 2003; Yantis e Serences, 2003; Figura 1). Várias tarefas top-down têm sido mostradas para envolver com sucesso regiões dorsais do córtex parietal, nomeadamente aquelas que envolvem espacial (Kastner et al., 1999; Corbetta et al., 2000; Hopfinger et al., 2000; Shomstein and Behrmann, 2006; Chiu and Yantis, 2009; Greenberg et al., 2010), bem como mudanças não espaciais de atenção (Giesbrecht et al., 2003; Yantis and Serences, 2003; Shomstein and Yantis, 2004b, 2006; Tamber-Rosenau et al., 2011).
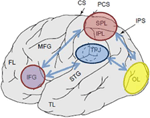
Figura 1. Representação esquemática de marcos anatômicos relevantes projetados na superfície lateral do cérebro humano. O lobo parietal Superior (SPL) e o lobo parietal inferior (IPL) são regiões dentro da parte dorsal do córtex parietal, preservando orientação de atenção top-down. A junção Temporo-parietal (TPJ) é uma região dentro do córtex parietal ventral que subsiste na orientação ascendente. Pensa-se que ambos, SPL e TPJ, induzem sinais de controle responsáveis pelas subsequentes modulações de atenção observadas sobre as regiões sensoriais, neste caso modulando (rotulado com setas azuis escuras) atividade evocada visualmente no lobo occipital (OL). Além disso, áreas ao longo do giro frontal inferior (IFG) e junção frontal inferior (IFJ) são consideradas como áreas de convergência para o controle de atenção impulsionado pelo estímulo e de cima para baixo (marcado por setas bidirecionais de azul claro).
em uma tarefa típica destinada a engajar a alocação de atenção de cima para baixo, os indivíduos são mostrados dois fluxos de apresentação visual em série rápida (RSVP) posicionados perifericamente e são inicialmente instruídos a monitorar um fluxo para um cue (por exemplo, um dígito entre o fluxo de letras). A identidade do cue indica se o sujeito deve manter a atenção no fluxo atual ou desviar a atenção para o outro fluxo (Yantis et al., 2002; Yantis and Serences, 2003). Dois grandes achados são observados em tais paradigmas. O primeiro tem a ver com o aumento da ativação dentro das regiões sensoriais que representam o local atendido no momento (por exemplo, aumento da atividade dentro das regiões visuais primárias da esquerda, quando o fluxo RSVP direito é atendido). Esta descoberta fornece provas sólidas de que os participantes estão a atender a um local específico e que a atenção modula a força da resposta sensorial (ver Figura 1; Moran e Desimone, 1985; O’Raven et al., 1997). A segunda constatação tem a ver com a observação de que as regiões dorsais do lobo parietal são selectivamente ativadas por turnos de atenção de cima para baixo. Observa-se que a duração da atividade do LPL/LPL é transitória, sugerindo que esta área do córtex parietal é a fonte de um breve sinal de controle de atenção para mudar os Estados atentos de uma maneira top-down (Yantis et al., 2002).vários estudos fMRI documentaram que a captura de atenção ascendente, mediada pela experiência e/ou relevância do estímulo, é preservada pela junção temporo-parietal (TPJ; Figura 1). Por exemplo, quando os sujeitos atendem e monitoram uma mudança em um estímulo visual ou auditivo, apresentado simultaneamente, a ativação das regiões TPJ do lobo parietal é aprimorada. Além da sensibilidade aparente a estímulos relevantes, o TPJ também é ativado em resposta a eventos potencialmente novos (inesperados ou pouco frequentes) Quando um organismo está envolvido em um contexto comportamental neutro ou quando envolvido em uma tarefa (Marois et al., 2000; Downar et al., 2002; Serences et al., 2005; Corbetta et al., 2008; Asplund et al., 2010; Diquattro e Geng, 2011; Geng e Mangun, 2011). Esta ativação ocorre independentemente da modalidade (auditiva, tátil e visual) em que a entrada é fornecida, refletindo a natureza multisensorial de TPJ (mas veja abaixo et al., 2001).
em uma tarefa típica examinando o mecanismo neural da captura de atenção bottom-up, os participantes são presenteados com um fluxo RSVP de itens no centro do visor e são convidados a identificar um alvo pré-definido (por exemplo, identificar a letra vermelha apresentada dentro de um fluxo RSVP de não-alvos brancos). Uma parte dos ensaios contém um distractor importante-tarefa irrelevante apresentado em vários intervalos de tempo antes do início do alvo, enquanto outros ensaios contêm apenas o distractor saliente (isto é, sem o alvo). Ensaios de “distractor-alvo” são utilizados para determinar a extensão da captura, mostrando que o distractor-tarefa irrelevante é, de facto, saliente, resultando assim numa diminuição da precisão do alvo. Os ensaios” distractor-in-isolation ” são utilizados para análises posteriores, uma vez que tais ensaios permitem o exame da actividade suscitada ao distractor saliente sem contaminação dos processos relacionados com o alvo. Vários achados importantes emergem de tais paradigmas. Em primeiro lugar, quando os distractores são espacialmente separados da localização alvo, os distractores de captura são acompanhados por uma maior actividade cortical nas regiões correspondentes do córtex sensorial (por exemplo, córtex visual retinotopicamente organizado; ver Figura 1). Tais resultados fornecem fortes provas de que, durante a captura, a atenção espacial é, de facto, captada para a localização espacial ocupada pelo distractor (Serences et al., 2005). Em segundo lugar, as regiões ventrais do córtex parietal, principalmente dentro do TPJ são ativadas seletivamente por mudanças de atenção involuntárias e bottom-up. Assim como a atividade dentro do SPL para a orientação top-down, o curso de atividade observado sobre TPJ é transitório na natureza sugerindo que esta região é a fonte de um breve sinal de controle de atenção para desviar a atenção de uma maneira bottom-up.
deve-se notar que enquanto esta revisão está focada no tratamento das funções cognitivas do córtex parietal posterior, outras regiões, notavelmente aquelas dentro do córtex frontal também são recrutadas para alocação de atenção top-down e bottom-up. Tais regiões incluem o córtex frontal ventral( VFC), os campos oculares frontais (FEF), junção frontal inferior (IFJ), e giro frontal inferior (IFG; Corbetta e Shulman, 2002, 2011; Serences et al., 2005; Asplund et al., 2010; Diquattro e Geng, 2011).
o papel do lobo Parietal na selecção de cima para baixo e de baixo para cima: A neuropsicologia
Historicamente, os pesquisadores invocado criticamente em estudos neuropsicológicos de pacientes com hemispatial negligência (um distúrbio de atribuição espacial de atenção para o esquerdo hemi-espaço) para obter informações sobre as funções cognitivas associadas com o lobo parietal. Na literatura neuropsicológica clássica, o córtex parietal, como um todo, foi geralmente considerado o local primário da lesão por negligência hemispacial. Esta visão, elaborada em detalhe pelos primeiros pesquisadores (Critchley, 1953; McFire e Zangwill, 1960; Piercy, 1964) reconheceu claramente a associação entre a lesão parietal e a negligência que se seguiu. Esta perspectiva foi em grande parte realizada através de 1980, quando Posner e colegas (1984) utilizou o covert visuo-espacial avançar paradigma para mostrar que os danos ao lobo parietal produz um déficit na “soltar” a operação (cancelamento de atenção a partir de um local e deslocando-a para o outro) quando o alvo é contralateral à lesão. No entanto, apesar deste grande avanço na compreensão da base neural da atenção e especificamente o papel de” desengate”do córtex parietal, seus achados assumem um único local cortical (córtex parietal) e uma única capacidade funcional (“desengate”). Em contraste com isso mais monolítico abordagem para o cérebro (córtex parietal) e o comportamento (atencional retirada), recente, comportamentais e de neuroimagem trabalho (acima analisados e em outros lugares) sugere que tanto a região cortical e as associadas atencional comportamento pode ser subdividido em qualitativamente diferentes perfis.
Dada a segregação das redes corticais no top-down e bottom-up de processos, uma óbvia previsão é de que os danos ao superior partes do parietal lobule (classificação SPL) deve produzir um déficit na meta-dirigida atencional, orientar, considerando que os danos para o inferior partes do parietal lobule (classificação TPJ) resultaria em um défice associado com o estímulo orientado a atenção de captura. Na medida em que estas correspondências de comportamento cerebral foram exploradas na literatura neuropsicológica, esta previsão não é obviamente confirmada. Por exemplo, os sintomas clínicos de negligência hemispacial estão fortemente associados a danos nas porções inferiores do lobo parietal, que inclui TPJ, ao invés de porções superiores como SPL (Friedrich et al., 1998; Shomstein et al., 2010; Corbetta e Shulman, 2011). Este é um pouco em desacordo com a neuroimagem literatura, o que sugere que o papel do TPJ está na captura da atenção, ao invés de incluir na orientação voluntária da atenção, o domínio em que a negligência pacientes parecem ter mais dificuldade. Para complicar ainda mais as coisas, observou-se que as lesões que envolvem SPL exclusivamente, só raramente produzem evidência clínica de negligência (Vallar e Perani, 1986). Outro estudo recente com pacientes com lesões centradas principalmente sobre TPJ e STG, mas preservado SPL, Corbetta et al. (2005) mostrou que a negligência espacial, bem como a sua recuperação, foi associada com a restauração da atividade tanto nas regiões parietais ventrais-parietais e dorsais (ver Corbetta e Shulman, 2011 para uma revisão). Embora interessante e empolgante em suas conclusões, este último estudo não diferencia a contribuição relativa das vias dorsal e ventral para diferentes tipos de atenção, uma vez que os pacientes foram testados apenas em uma variante da tarefa de avaliação de atenção espacial Posner, tarefa que se acredita envolver tanto de cima para baixo quanto de baixo para cima.para distinguir entre controle de atenção orientado por objetivos e captura de atenção saliente e para examinar seu mapeamento no SPL e TPJ, respectivamente, estudo recente adotou dois paradigmas comportamentais, cada um visando uma dessas formas de atenção (Shomstein et al., 2010). Para examinar a integridade da orientação de atenção top-down nos pacientes, uma tarefa top-down foi usada exigindo que os participantes mudassem a atenção espacial entre as correntes RSVP separadas espacialmente (uma tarefa que tem sido usada com sucesso para demonstrar ativação SPL em estudos fMRI (Yantis et al., 2002)). Da mesma forma, a fim de examinar as habilidades de orientação de atenção de baixo para cima dos pacientes, uma variante de Folk et al. (2002) contingent capture paradigm was employed in which participants detected targets that appeared at fixation while task-irrelevant color singletons were flashed in the periphery. A medida em que os distractores da tarefa-irrelevantes interferem com a tarefa central de detecção foi então usada como uma medida de captura de atenção ascendente (Bacon e Egeth, 1994; Folk et al., 2002).as previsões foram as seguintes:: pacientes com lesões superior partes do lobo parietal (que afectam SPL) deve ser prejudicada no top-down atencional orientação de tarefas (com desempenho preservado na captura de tarefas), enquanto os pacientes com lesões de partes inferiores do lobo parietal (que afectam TPJ) deve ser prejudicada na captura de tarefas (com poupado de desempenho no top-down tarefa). Uma dupla dissociação desta forma não só atesta os componentes independentes da atenção, mas também sugere que tais componentes de atenção são mediados por mecanismos neurais independentes. Oito pacientes com negligência visual-espacial foram recrutados para o estudo e completaram duas tarefas, batendo em orientação orientada a estímulos ou orientada a objetivos. Com base em seu perfil comportamental, os pacientes foram classificados em grupos e sua sobreposição de lesão foi explorada (figura 2A). Pacientes que exibiram dificuldades com orientação orientada para o objetivo, como quantificado pelo Índice de atenção top-down (figura 2B), apresentaram sobreposição da lesão centrada em porções superiores do lóbulo parietal (SPL subsumente) com o lóbulo parietal inferior poupado (TPJ). Pacientes com lesão sobrepõem-se centrados sobre as porções inferiores do lobo parietal (subumagem TPJ), mas pouparam SPL realizada normalmente na tarefa de orientação dirigida para o objetivo, enquanto permanecem imunes à captura de atenção (figura 2C). Os resultados deste estudo sugerem claramente que o SPL e o TPJ são regiões anatômicas que são necessariamente recrutadas para fins de orientação de cima para baixo e de baixo para cima e que os danos ao SPL e ao TPJ levam a desordens de orientação de cima para baixo e de baixo para cima, respectivamente.
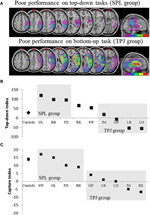
Figura 2. Resultados do estudo neuropsicológico destinado a investigar a contribuição relativa do SPL e do TPJ para a orientação de cima para baixo e de baixo para cima. (A) Lesão sobreposições (roxo sobreposição mínima; vermelho máxima sobreposição) para pacientes agrupados por déficits comportamentais no top-down atencional, orientar, rotulado como o SPL grupo (painel superior); e pacientes agrupados por déficits comportamentais em bottom-up, orientar, rotulado a TPJ grupo (painel inferior). (B) Performance comportamental na tarefa de cima para baixo resumida com um “índice de cima para baixo” que quantifica as diferenças entre as mudanças espaciais de cima para baixo feitas da esquerda para a direita e vice-versa. Controles e o grupo TPJ lesioned mostram eficiência similar na execução de mudanças espaciais, enquanto pacientes com lesões SPL mostram menor eficiência. O controlo do grupo e os dados individuais do doente (marcados com as iniciais do doente) são desenhados na abcissa. (C)” Índice de captura ” é uma medida da atenção ascendente e quantifica a medida em que os distractores irrelevantes capturam a atenção para longe da tarefa. Controles e o grupo SPL lesioned mostram valores de captura similares, de modo que ambos os grupos são capturados pelos distractores da tarefa-irrelevantes. O grupo TPJ lesioned mostra Índice de captura muito reduzido (falha em ser capturado). Note-se que os pacientes foram colocados no grupo SPL ou TPJ com base no comportamento, ao invés de com base na lesão, notando assim a consistência com que os pacientes acabam no grupo correspondente.
interacção entre a selecção de cima para baixo e de baixo para cima
embora aparentemente exista uma forte associação entre a orientação orientada para o objectivo e a SPL e a orientação orientada para o estímulo e TPJ, dados de Shomstein et al. (2010) estudo de doentes sugere que estes dois sistemas não são totalmente independentes. Esta conclusão é apoiada pela constatação de que os pacientes com dano SPL exibiram um padrão de desempenho rotulado como “captura hiper”.”Ao contrário dos controles, para os quais apenas o distractor alvo colorido captou a atenção (levando a menor precisão do alvo), distractores coloridos irrelevantes também provaram estar distraindo para pacientes com lesão SPL. Além disso, enquanto para os controles a atenção foi capturada por distractores apenas quando eles precederam o início do alvo, para pacientes com lesões SPL a atenção foi mesmo capturada por distractores apresentados simultaneamente com o alvo. Este padrão de desempenho pode ser explicado pelo seguinte quadro:: O SPL é responsável pela orientação top-down da atenção que inclui a determinação dos aspectos dos estímulos que são relevantes para a tarefa (por exemplo, Busca pelo alvo vermelho; Corbetta e Shulman, 2002; Serences et al., 2005). Este conjunto de atenção então restringe o TPJ, de tal forma que a captura do mecanismo de atenção que é mediado pelo TPJ é apenas desencadeada pela informação relevante da tarefa (por exemplo, distractores vermelhos capturando a atenção, e distractores cinzentos não capturando a atenção ao procurar um alvo vermelho). A ausência de SPL impede o estabelecimento de um conjunto de atenção relevante para uma tarefa e, portanto, qualquer estímulo, tarefa relevante ou não, é considerado importante, portanto, captar a atenção (por exemplo, distractor tarefa-irrelevante que captura a atenção para o grupo SPL) indiscriminadamente.
tem sido sugerido que SPL e TPJ poderiam interagir de pelo menos uma de duas maneiras possíveis. A primeira possibilidade é que o TPJ serve como um sistema de alerta que detecta estímulos comportamentalmente relevantes, mas não tem a alta resolução espacial, assim quando um estímulo comportamentalmente relevante é detectado sua localização precisa é fornecida pelo SPL que armazena mapas espaciais (Kastner et al., 1999; Wojciulik and Kanwisher, 1999; Bisley and Goldberg, 2003; Silver et al., 2005). Uma possibilidade hipotética relacionada é que o mecanismo de captura (que inclui TPJ) atua como um disjuntor de atividade cognitiva contínua quando um estímulo relevante comportamentalmente é apresentado (Corbetta e Shulman, 2002, 2011). O “hiper-captura” padrão de atividade observado em pacientes com preservado TPJ, mas lesioned SPL fornece mais evidências para a hipótese de que vistas TPJ como a emissão de um sinal de controle que termina a tarefa à mão, assim, servir como um disjuntor (Corbetta e Shulman, 2002; Serences et al., 2005). Outros estudos recentes de neuroimagem que empregam vários paradigmas forneceram mais evidências para uma relação interativa entre a orientação de cima para baixo e a orientação de baixo para cima, e, posteriormente, para a relação entre SPL e TPJ (Serences et al., 2005; Asplund et al., 2010; Diquattro e Geng, 2011).embora a evidência de uma interação entre os dois sistemas de atenção e os dois substratos de atenção (SPL e TPJ) seja forte, o que permanece incerto é se esta interação é direta entre SPL e TPJ ou se é realizada através de outras regiões intermediárias. Como foi mencionado anteriormente, as redes de orientação de atenção de cima para baixo e de baixo para cima envolvem várias regiões dentro do córtex frontal, portanto, é razoável supor que a convergência entre os dois sistemas pode ser realizada através do lobo frontal. Dois estudos recentes que investigaram a interacção entre a selecção de atenção de cima para baixo e de baixo para cima forneceram provas para o IFJ e o IFG como possíveis locais de convergência entre a selecção orientada por estímulos e a selecção orientada por objectivos (Asplund et al., 2010; Diquattro e Geng, 2011). O IFJ e o IFG parecem ser candidatos ideais para tal interação, dado o seu envolvimento geral na atenção e no controle cognitivo, bem como o seu envolvimento na seleção espacial e não espacial (Koechlin et al., 2003; Brass et al., 2005).
o papel do Lobo Parietal na selecção de cima para baixo e de baixo para cima: Fisiologia
embora a ênfase desta revisão tenha sido predominantemente colocada em estudos humanos, uma grande riqueza de conhecimentos sobre o envolvimento do córtex parietal na orientação de atenção foi colhida a partir de investigações de fisiologia de macacos (ver revisão recente de Bisley e Goldberg, 2010). No entanto, quando se trata de examinar as contribuições relativas de diferentes regiões dentro do córtex parietal para orientação de cima para baixo e de baixo para cima, a literatura fisiológica do Macaco fica aquém. A principal razão para isso é que dentro do córtex do macaco não parece haver evidência para a mesma segregação do controle de cima para baixo e de baixo para cima. Em vez disso, o lateral intraparietal área (LIP) originalmente pensado para ser envolvido na sacca de planejamento (Gnadt e Andersen, 1988) está envolvido na atenção visual e funciona como um mapa de prioridades em que estímulos externos são representados de acordo com a sua comportamental prioridade derivada no top-down ou bottom-up maneira (Colby e Goldberg, 1999; Bisley e Goldberg, 2003, 2010; Balan e Gottlieb, 2006; Ipata et al., 2006; Buschman and Miller, 2007; Gottlieb and Balan, 2010).
Conclusão
Embora, muito menos é conhecido sobre o córtex parietal do que homólogo macaco córtex, estudos recentes, empregando neuroimagem e neuropsicológicos métodos, começaram a elucidar cada vez mais refinadas funcionais e estruturais distinções. Esta revisão centrou-se em estudos neuroimaging e neuropsicológicos recentes que elucidam os papéis cognitivos das regiões dorsais e ventrais do córtex parietal na orientação de atenção de cima para baixo e de baixo para cima, e na interação entre os dois mecanismos de alocação de atenção.
Declaração de conflito de interesses
o autor declara que a investigação foi realizada na ausência de quaisquer relações comerciais ou financeiras que possam ser interpretadas como um potencial conflito de interesses.Este trabalho foi apoiado pelo National Institutes of Health grant EY021644.
Asplund, C. L., Todd, J. J., Snyder, A. P., and Marois, R. (2010). Um papel central para o córtex pré-frontal lateral na atenção orientada para o objetivo e impulsionada pelo estímulo. Conversao. Neuroci. 13, 507–512.
Pubmed Abstract | Pubmed Full Text / CrossRef Full Text
Bacon, W. F., And Egeth, H. E. (1994). Captura convencional impulsionada pelo estímulo. Percept. Psychophys. 55, 485–496.
Pubmed Abstract / Pubmed texto completo
Balan, P. F., and Gottlieb, J. (2006). Integração da contribuição exógena num mapa dinâmico de saliências revelado pela atenção perturbadora. J. Neurosci. 26, 9239–9249.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Behrmann, M., Geng, J. J., and Shomstein, S. (2004). Córtex Parietal e atenção. Moeda. Opin. Neurobiol. 14, 212–217.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Bisley, J. W., and Goldberg, M. E. (2003). Actividade Neuronal na área intraparietal lateral e atenção espacial. Science 299, 81-86.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Bisley, J. W., and Goldberg, M. E. (2010). Atenção, intenção e prioridade no lobo parietal. Annu. Rev. Neurosci. 33, 1–21.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Brass, M., Derrfuss, J., Forstmann, B., and von Cramon, D. Y. (2005). O papel da área de junção frontal inferior no controle cognitivo. Trends Cogn. Ciência. 9, 314–316.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Buschman, T. J., and Miller, E. K. (2007). Controle de cima para baixo versus baixo para cima da atenção nos vértices pré-frontal e posterior parietal. Science 315, 1860-1862.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Chiu, Y. C., and Yantis, S. (2009). Uma fonte independente de domínio de controle cognitivo para conjuntos de Tarefas: mudança de atenção espacial e regras de categorização de comutação. J. Neurosci. 29, 3930–3938.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Colby, C. L., and Goldberg, M. E. (1999). Espaço e atenção no córtex parietal. Annu. Rev. Neurosci. 22, 319–349.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., Kincade, J. M., Ollinger, J. M., McAvoy, M. P., and Shulman, G. L. (2000). A orientação voluntária é dissociada da detecção de alvos no córtex parietal posterior humano. Conversao. Neuroci. 3, 292–297.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., Kincade, M. J., Lewis, C., Snyder, A. Z., and Sapir, A. (2005). Base Neural e recuperação de déficits de atenção espacial em negligência espacial. Conversao. Neuroci. 8, 1603–1610.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., Patel, G., and Shulman, G. L. (2008). The reorienting system of the human brain: from environment to theory of mind. Neuron 58, 306-324.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Corbetta, M., and Shulman, G. L. (2002). Controlo da atenção dirigida ao objectivo e impulsionada pelo estímulo no cérebro. Conversao. Rev. Neurosci. 3, 201–215.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., and Shulman, G. L. (2011). Redes de negligência espacial e de atenção. Annu. Rev. Neurosci. 34, 569–599.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Critchley, M. (1953). parietal. London, UK: Hafner Press.
Pubmed Abstract/Pubmed Full Text
Diquattro, N. E., and Geng, J. J. (2011). O conhecimento Contextual configura redes de controlo de atenção. J. Neurosci. 31, 18026–18035.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Downar, J., Crawley, A. P., Mikulis, D. J., and Davis, K. D. (2001). The effect of task relevance on the cortical response to changes in visual and auditory stimuli: an event-related fMRI study. Neuroimagem 14, 1256-1267.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Downar, J., Crawley, A. P., Mikulis, D. J., and Davis, K. D. (2002). Uma rede cortical sensível à experiência do estímulo num contexto comportamental neutro através de múltiplas modalidades sensoriais. J. Neurofisiol. 87, 615–620.
Pubmed Abstract / Pubmed Full Text
Drummond, L., and Shomstein, S. (2010). Atenção baseada em objectos: mudança ou incerteza? Atten. Percept. Psychophys. 72, 1743–1755.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Egeth, H. E., and Yantis, S. (1997). Atenção Visual: controle, representação e curso de tempo. Annu. Rev. Psychol. 48, 269–297.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Eriksen, C. W., and Hoffman, J. E. (1972). Características temporais e espaciais da codificação selectiva a partir de ecrãs visuais. Percept. Psychophys. 12, 201–204.
Folk, C. L., Leber, A. B., and Egeth, H. E. (2002). Fez-te piscar os olhos! Captura de atenção contingente produz um piscar de olhos espacial. Percept. Psychophys. 64, 741–753.
Pubmed Abstract / Pubmed Full Text
Folk, C. L., Remington, R. W., and Johnston, J. C. (1992). Orientação secreta involuntária depende das configurações de controlo de atenção. J. Exp. Psychol. Cantarolar. Percept. Executar. 18, 1030–1044.
Pubmed Abstract/Pubmed Full Text
Friedrich, F. J., Egly, R., Rafal, R. D., and Beck, D. (1998). Spatial attention deficits in humans: a comparison of superior parietal and temporal-parietal junction lesions. Neuropsicologia 12, 193-207.
Pubmed Abstract / Pubmed Full Text
Geng, J. J., and Behrmann, M. (2002). A avaliação de probabilidade da localização do alvo facilita a busca visual implicitamente em participantes normais e pacientes com negligência hemispacial. Psychol. ciência. 13, 520–525.
Pubmed Abstract / Pubmed Full Text
Geng, J. J., and Behrmann, M. (2005). Probabilidade espacial como um sinal de atenção na busca visual. Percept. Psychophys. 67, 1252–1268.
Pubmed Abstract / Pubmed Full Text
Geng, J. J., and Mangun, G. R. (2011). A ativação da junção temporoparietal direita por uma sugestão contextual saliente facilita a discriminação do alvo. Neuroimagem 54, 594-601.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Gibson, B. S., and Kelsey, E. M. (1998). A captura de atenção impulsionada pelo estímulo depende de um conjunto de atenção para as características visuais de todo o display. J. Exp. Psychol. Cantarolar. Percept. Executar. 24, 699–706.
Pubmed Abstract / Pubmed Full Text
Giesbrecht, B., Woldorff, M. G., Song, A. W., and Mangun, G. R. (2003). Mecanismos neurais de controle de cima para baixo durante a atenção espacial e característica. Neuroimagem 19, 496-512.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Gnadt, J. W., and Andersen, R. A. (1988). Actividade de planeamento motor relacionada com a memória no córtex parietal posterior do macaco. Expo. Brain Res. 70, 216-220.
Pubmed Abstract / Pubmed Full Text
Gottlieb, J., and Balan, P. (2010). Atenção como uma decisão no espaço de informação. Trends Cogn. Ciência. 14, 240–248.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Greenberg, A. S., Esterman, M., Wilson, D., Serences, J. T., and Yantis, S. (2010). Controle de atenção espacial e baseada em recursos no córtex frontoparietal. J. Neurosci. 30, 14330–14339.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Hopfinger, J. B., Buonocore, M. H., and Mangun, G. R. (2000). Os mecanismos neurais do controlo de atenção de cima para baixo. Conversao. Neuroci. 3, 284–291.
Pubmed Abstract | Pubmed texto completo / CrossRef texto completo
Ipata, A. E., Caramba, A. L., Gottlieb, J., Bisley, J. W., e Goldberg, M. E. (2006). As respostas labiais a um estímulo popout são reduzidas se forem ignoradas abertamente. Conversao. Neuroci. 9, 1071–1076.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Johnston, W. A., and Dark, V. J. (1986). Atenção Selectiva. Annu. Rev. Psychol. 37, 43–75.Juola, J. F., Koshino, H., and Warner, C. B. (1995). Trocas entre efeitos de atenção de pistas espaciais e onsets abruptos. Percept. Psychophys. 57, 333–342.
Pubmed Abstract / Pubmed Full Text
Kastner, S., Pinsk, M. A., de Weerd, P., Desimone, R., and Ungerleider, L. G. (1999). Aumento da actividade no córtex visual humano durante a atenção dirigida na ausência de estimulação visual. Neuron 22, 751-761.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Koechlin, E., Ody, C., and Kouneiher, F. (2003). A arquitetura do controle cognitivo no córtex pré-frontal humano. Science 302, 1181-1185.
Pubmed Resumo | Pubmed Texto Completo | CrossRef Texto Completo
Koshino, H.), Warner, C. B., and Juola, J. F. (1992). Eficácia relativa de sinais centrais, periféricos e de início abrupto na atenção visual. Q. J. Exp. Psychol. A 45, 609-631.
Pubmed Abstract / Pubmed Full Text
Liu, T., Slotnick, S. D., Serences, J. T., and Yantis, S. (2003). Mecanismos corticais de controlo de atenção baseado em recursos. Cérebro. Cortex 13, 1334-1343.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Marois, R., Leung, H. C., and Gore, J. C. (2000). Uma abordagem motivada pelo estímulo à identidade do objecto e ao processamento da localização no cérebro humano. Neuron 25, 717-728.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
McFire, J., and Zangwill, O. L. (1960). Deficiências visuais-construtivas associadas a lesões do hemisfério cerebral esquerdo. Cérebro 82, 243-260.
Moore, C. M., and Egeth, H. (1998). Como a atenção baseada em recursos afeta o processamento visual? J. Exp. Psychol. Cantarolar. Percept. Executar. 24, 1296–1310.
Pubmed Abstract / Pubmed texto completo
Moran, J., and Desimone, R. (1985). Portas de atenção selectivas processamento visual no córtex extra-triado. Science 229, 782-784.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
O’Craven, K., Rosen, B. R., Kwong, K. K., Treisman, A., and Savoy, R. L. (1997). A atenção voluntária modula a actividade da fMRI no MT-MST humano. Neuron 18, 591-598.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Piercy, M. (1964). The effects of cerebral lesions on intellectual function: a review of current research trends. Br. J. Psychiatry 110, 310-352.
Pubmed Abstract / Pubmed texto completo
Posner, M. I. (1980). Orientação da atenção. Q. J. Exp. Psychol. 32, 3–25.
Pubmed Abstract / Pubmed Full Text
Posner, M. I., Snyder, C. R., and Davidson, B. J. (1980). Atenção e detecção de sinais. J. Exp. Psychol. 109, 160–174.
Pubmed Abstract / Pubmed Full Text
Posner, M. I., Walker, J. A., Friedrich, F. J., and Rafal, R. D. (1984). Efeitos da lesão parietal na orientação secreta da atenção. J. Neurosci. 4, 1863–1874.
Pubmed Abstract/Pubmed Full Text
Serences, J. T., Shomstein, S., Leber, A. B., Golay, X., Egeth, H. E., and Yantis, S. (2005). Coordenação do controlo de atenção voluntário e orientado para o estímulo no córtex humano. Psychol. Ciência. 16, 114–122.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Shaw, M. L. (1978). Um modelo de atribuição de capacidade para o tempo de reacção. J. Exp. Psychol. Cantarolar. Percept. Executar. 4, 586–598.
Shomstein, S., and Behrmann, M. (2006). Sistemas corticais que mediam a atenção visual tanto para objectos como para localizações espaciais. Procedimento. Natl. Acad. Ciência. U. S. A. 103, 11387-11392.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Shomstein, S., Lee, J., and Behrmann, M. (2010). Orientação de atenção Top-down e bottom-up: investigating the role of the dorsal and ventral parietal cortices. Expo. Brain Res. 206, 197-208.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Shomstein, S., and Yantis, S. (2004a). Priorização configurural e contextual na atenção baseada em objetos. Psychon. Touro. Rev. 11, 247-253.
Pubmed Abstract / Pubmed Full Text
Shomstein, S., and Yantis, S. (2004b). O controle da atenção muda entre visão e audição no córtex humano. J. Neurosci. 24, 10702–10706.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Shomstein, S., and Yantis, S. (2006). O córtex Parietal medeia o controle voluntário da atenção auditiva espacial e não espacial. J. Neurosci. 26, 435–439.
Pubmed Abstract | Pubmed texto completo / CrossRef texto completo
Silver, M. A., Ress, D., and Heeger, D. J. (2005). Mapas topográficos de atenção espacial visual no córtex parietal humano. J. Neurofisiol. 94, 1358–1371.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Tamber-Rosenau, B. J., Esterman, M., Chiu, Y.-C., and Yantis, S. (2011). Mecanismos corticais de controle cognitivo para a mudança de atenção na visão e memória de trabalho. J. Cogn. Neuroci. 23, 2905–2919.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Theeuwes, J. (1991). Controlo exógeno e endógeno da atenção: o efeito de onsets visuais e compensações. Percept. Psychophys. 49, 83–90.
Pubmed Abstract / Pubmed Full Text
Vallar, G., and Perani, D. (1986). A anatomia de negligência unilateral após lesões de AVC no hemisfério direito. Um estudo de correlação clínica/TC-scan no homem. Neuropsicologia 24, 609-622.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Wojciulik, E., and Kanwisher, N. (1999). A generalidade do envolvimento parietal na atenção visual. Neuron 23, 747-764.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Yantis, S., and Jonides, J. (1984). Sinais visuais abruptos e atenção seletiva: evidência de busca visual. J. Exp. Psychol. Cantarolar. Percept. Executar. 10, 601–621.
Pubmed Abstract / Pubmed Full Text
Yantis, S., and Jonides, J. (1990). Movimentos visuais bruscos e atenção selectiva: distribuição voluntária versus automática. J. Exp. Psychol. Cantarolar. Percept. Executar. 16, 121–134.
Pubmed Abstract / Pubmed Full Text
Yantis, S., Schwarzbach, J., Serences, J. T., Carlson, R. L., Steinmetz, M. A., Pekar, J. J., e Courtney, S. M. (2002). Atividade neural transitória no córtex parietal humano durante mudanças de atenção espacial. Conversao. Neuroci. 5, 995–1002.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Yantis, S., and Serences, J. T. (2003). Mecanismos corticais de controlo de atenção baseado no espaço e em objetos. Moeda. Opin. Neurobiol. 13, 187–193.
Pubmed Abstract / Pubmed texto completo / CrossRef texto completo