Introduction
Fossil resources are the main feedstock for energy and organic compounds, and their use results in the emission of greenhouse gases associated with climate change. O próximo crash climático exige uma transição urgente de uma economia baseada em fósseis para uma economia baseada em bio-base na qual a biomassa lignocelulósica em vez de petróleo é usado para a produção de combustíveis, produtos químicos e materiais. A madeira é uma importante fonte de biomassa lignocelulósica; é constituído principalmente por paredes celulares de espessura secundária ricas em celulose, hemiceluloses e lignina. Todos os três polímeros podem ser valorizados na economia bio-baseada. A celulose é uma fonte para a indústria de celulose e papel, e tanto a celulose e hemiceluloses podem ser despolimerizadas aos seus monossacáridos para fermentação em, por exemplo, bio-etanol, ácido láctico e detergentes (Vanholme et al., 2013b). Como a lignina afeta negativamente a eficiência do processamento de madeira para essas aplicações, as árvores podem ser projetadas para acumular menos lignina, para se tornar mais amenizável para a produção de papel e açúcares fermentáveis. Por outro lado, a lignina é cada vez mais considerada uma componente valiosa na economia baseada na biotecnologia. Com efeito, dado que a lignina é a maior fonte aromática renovável na terra, a viabilidade económica de uma bio-refinaria pode ser significativamente aumentada se a lignina também for valorizada e utilizada como recurso para a produção de produtos químicos (Holladay et al., 2007; Tuck et al., 2012; Davis et al., 2013; Ragauskas et al., 2014; Li C. et al., 2015; Van den Bosch et al., 2015; Rinaldi et al., 2016; Upton and Kasko, 2016; Schutyser et al., 2018).
O polímero de lignina é composto por monolignóis produzidos pelas vias biossintéticas fenilpropanóide e monolignol, por uma série de reacções enzimáticas começando com a desaminação da fenilalanina (Figura 1). Os monolignóis são sintetizados no citoplasma e translocados para o apoplasto, onde são desidrogenados a Radicais monolignol pela ação de lacases e peroxidases (Berthet et al., 2011; Zhao et al., 2013). Estes monolignol radicais, em seguida, casal com os outros em uma combinatória de moda, gerando uma série de ligações químicas, tais como o teluretos aromáticos-éter bond (β-O-4), resinol bond (β-β), e phenylcoumaran bond (β-5) (Boerjan et al., 2003; Ralph et al., 2004; Vanholme et al., 2010). Os monolignóis mais comuns são os álcoois p-cumaril, coniferil e sinapil, que geram as Unidades H, G E S após sua incorporação no polímero de lignina, respectivamente (Bonawitz e Chapple, 2010; Ralph et al., 2019; Vanholme et al., 2019). A contribuição relativa dos blocos de construção de lignina varia entre taxa, fase de desenvolvimento, tipo de tecido e célula, e até mesmo camada de parede celular; Lignina de madeiras macias (gymnosperms) é composta quase inteiramente de unidades G com uma pequena fração de Unidades H, enquanto Lignina de madeiras duras (angiospérmicas) tem unidades S além de unidades G e traços de Unidades H (Boerjan et al., 2003; Vanholme et al., 2010, 2019). Além destes monolignóis tradicionais, uma variedade de outras moléculas aromáticas p-hidroxiladas pode ser incorporada no polímero de lignina a vários níveis (Vanholme et al., 2019).
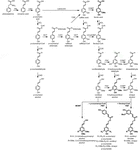
Figure 1. Lignin biosynthetic pathway. Alternative monomers and heterologously expressed enzymes are shown in bold. PAL, phenylalanine ammonia-lyase; C4H, cinnamate 4-hydroxylase; 4CL, 4-coumarate:CoA ligase; HCT, p-Hydroxycinnamoyl-CoA:quinate/shikimate-p-hydroxycinnamoyltransferase; C3’H, p-coumaroyl quinate/shikimate 3’-hydroxylase; CSE, caffeoyl shikimate esterase; CCoAOMT, caffeoyl-CoA O-methyltransferase; CCR, cinnamoyl-CoA reductase; F5H, ferulate 5-hydroxylase/CAld5H, coniferaldehyde 5-hydroxylase; COMT, ácido cafeico o-metiltransferase; CAD, álcool cinamílico desidrogenase; FMT, feruloil-CoA monolignol transferase; PMT, p-cumaroil-CoA monolignol transferase.
uma vez que a lignina é um dos principais recalcitrance fator de madeira na deslignificação processos, grandes esforços de pesquisa têm se dedicado a desvendar a lignina biossintéticos caminho, e para estudar os efeitos das perturbações da biossíntese de lignina genes de lignina, a quantidade e a composição, e no processamento de madeira da eficiência. Enquanto modificações em genes que vão desde os fatores de transcrição codificados até aqueles que codificam enzimas oxidativas resultaram em conteúdo alterado de lignina, composição ou deposição (Eriksson et al., 2000; Li Y. H. et al., 2003; Liang et al., 2008; Lu et al., 2013; Lin et al., 2016; Xu et al., 2017; Yang et al., 2017; Obudulu et al., 2018), esta revisão incidirá sobre os resultados obtidos pela engenharia dos genes biossintéticos da lignina.
Engenharia da Via da lignina
a Tabela 1 fornece uma visão geral dos diferentes estudos sobre genes biossintéticos da lignina em baixa regulação ou mutação em choupo, pinheiro, eucalipto e bétula, com os efeitos resultantes na eficiência de processamento da madeira, quando determinada. A redução da actividade de qualquer etapa da via biossintética da lignina, desde PAL até CAD, pode resultar numa redução do teor de lignina (Tabela 1). Vários parâmetros influenciam o grau de redução de lignina, tais como o gene alvo e o grau de downregulation da atividade da enzima, que por sua vez depende da eficiência do silêncio construção usada, o tamanho da família do gene, e a redundância dentro da família do gene. De um modo geral, a redução das etapas de C4H até CCR resulta numa redução mais drástica da quantidade de lignina (Hu et al., 1999; Meyermans et al., 2000; Zhong et al., 2000; Li L. et al., 2003; Jia et al., 2004; Lu et al., 2004; Leplé et al., 2007; Coleman et al.,2008a, b; Bjurhager et al., 2010; Mansfield et al., 2012; Ralph et al., 2012; Min et al., 2014; Van Acker et al., 2014; Zhou et al., 2015, 2018; Saleme et al., 2017; Xiang et al., 2017) do que o regulamento de redução de F5H, COMT e CAD (Van Doorsselaere et al., 1995; Baucher et al., 1996; Lapierre et al., 1999; Jouanin et al., 2000; Van Acker et al., 2014; Wang et al., 2018). A redução da lignina pode ser associada a um aumento em S / G, Como EM C3 ‘ h- (Coleman et al., 2008a; Ralph et al., 2012) and CCoAOMT-downregulated trees (Meyermans et al., 2000), ou uma diminuição na razão S / G, tal como na CSE- (Saleme et al., 2017), and COMT-downregulated trees (Van Doorsselaere et al., 1995; Lapierre et al., 1999; Jouanin et al., 2000). Curiosamente, verificou-se que os poplares de lignina 4CL com baixo teor de enxofre têm um aumento de S/G (Min et al., 2014; Xiang et al., 2017), uma diminuição em S / G (Voelker et al., 2010; Zhou et al., 2015), ou rácios comparáveis ao tipo selvagem (Hu et al., 1999; Li L. et al., 2003). Esta variação não pode ser associada ao promotor ou ao método utilizado para a redução da regulamentação, sugerindo que as diferenças no grau de silenciamento, condições de crescimento ou estado de desenvolvimento influenciam Esta característica. Por outro lado, observaram-se os efeitos mais fortes sobre a composição de lignina H/G/S para as árvores que foram objecto de uma baixa regulamentação em C3 ‘ h E HCT, que depositam lignina com grandes aumentos do conteúdo da unidade H (Coleman et al., 2008a; Ralph et al., 2012; Vanholme et al., 2013a), enquanto as árvores que sobreexpressam F5H produzem lignina fortemente enriquecida em unidades S (Franke et al., 2000; Li L. et al., 2003; Stewart et al., 2009), e as árvores que estão em baixa regulamentação na COMT reduziram drasticamente a biossíntese da unidade S (Van Doorsselaere et al., 1995; Lapierre et al., 1999).

a Tabela 1. Visão geral das árvores florestais com expressão modificada de genes de biossíntese de lignina.
A eficiência de processamento da biomassa também pode ser modificada pelo aumento da incorporação de moléculas que representam, geralmente, menor componentes na lignina do tipo selvagem plantas. A incorporação de ácido ferúlico em árvores deficientes em CCR resulta na formação de ligações acetais no polímero de lignina, que são facilmente clivadas em pré-tratamentos de biomassa ácida (Leplé et al., 2007; Ralph et al., 2008; Van Acker et al., 2014). De facto, os níveis de ácido ferúlico na lignina correlacionaram-se positivamente com uma maior eficiência de sacarificação (Van Acker et al., 2014). A incorporação de álcool 5-hidroxiconiferílico e 5-hidroxiconiferaldeído na Lignina de poplares deficientes em COMT (Van Doorsselaere et al., 1995; Lapierre et al., 1999; Jouanin et al., 2000; Morreel et al., 2004; Lu et al., 2010) dá origem a ligações benzodioxanas, impedindo potencialmente a ligação covalente entre a lignina e os grupos hidroxila polissacáridos (Weng et al., 2010; Vanholme et al., 2012; Nishimura et al., 2018). Por outro lado, a deficiência da COMT resulta também numa lignina mais condensada devido aos níveis relativamente mais elevados das ligações β-β e β-5 condensadas e aos níveis mais baixos das ligações β-o-4, Quando a frequência da unidade S diminui. A polpa química de madeira derivada de poplares fortemente desregulados para a COMT resultou numa maior produção de pasta, contrabalançada pelo teor residual de lignina na pasta. Estas árvores tinham uma lignina mais baixa e um teor de celulose mais elevado (Jouanin et al., 2000). Por outro lado, os poplares que foram modestamente reduzidos para a COMT tiveram uma grande diminuição no rendimento da pasta de papel, presumivelmente porque o teor de lignina tinha permanecido normal, enquanto a lignina tinha uma maior frequência de ligações condensadas que afetaram negativamente a extração de lignina (Lapierre et al., 1999; Pilatos et al., 2002). A incorporação de cinamaldehydes no polímero de lignina em árvores deficientes em CAD resulta em cadeias de polímeros de lignina mais curtas, daí uma maior proporção de grupos terminais fenólicos livres que aumentam a solubilidade do polímero em álcali. A incorporação de cinamaldehydes no polímero de lignina provavelmente também reduz a interação covalente da cadeia alifática com hemicelulose, tornando novamente a lignina mais solúvel. Além disso, devido ao sistema conjugado prolongado que é gerado quando UM β-o-4 de cinamaldeído casais com outro monômero, a ligação de éter aromático do cinamaldeído incorporado torna-se mais suscetível à clivagem alcalina (Lapierre et al., 1989; Van Acker et al., 2017).
polimerização de lignina é um processo de acoplamento RADical combinatório, permitindo que uma ampla gama de compostos fenólicos sejam naturalmente incorporados no polímero de lignina (Boerjan et al., 2003; Vanholme et al., 2019). Pesquisadores tentaram adaptar a quantidade e composição de lignina para melhorar o processamento de biomassa pela expressão de genes heterólogos, visando a biossíntese e incorporação de vários compostos fenólicos compatíveis como monolignóis alternativos no polímero de lignina (Ralph, 2006; Vanholme et al., 2012; Mottiar et al., 2016; Mahon e Mansfield, 2018). Um exemplo é a introdução de genes que codificam enzimas que são necessárias para a biossíntese da unidade S no pinho; a expressão simultânea de F5H, COMT e CAD introduziu com sucesso unidades S em Pinus radiata (Wagner et al., 2015; Edmunds et al., 2017). A introdução do gene que codifica um monolignol 4-o-metiltransferase (MOMT4) em poplar leva à formação de álcoois 4-o-metilados coniferil e sinapil, que não podem ser incorporados no crescente polímero de lignina por falta do grupo hidroxilo aromático. Isso leva a uma parada na polimerização da lignina e resulta em árvores com menor conteúdo de lignina e maior eficiência de sacarificação (Bhuiya e Liu, 2010; Cai et al., 2016). Poplars também foram projetados para conter ligações ester na espinha dorsal de polímero de lignina. Os ésteres ferulados de coniferilo foram introduzidos no polímero através da expressão de um FERULOIL-CoA:gene da MONOLIGNOL TRANSFERASE (FMT) derivado de Angelica sinensis (Wilkerson et al., 2014), levando a uma maior eficiência de sacarificação sob várias condições de pré-tratamento (Wilkerson et al., 2014; Kim et al., 2017; Bhalla et al., 2018), e uma maior eficiência de pulverização de Kraft em comparação com o tipo selvagem (Zhou et al., 2017). Monolignol p-coumarate ésteres também foram projetados em poplar, através da expressão de um arroz p-COUMAROYL-CoA:MONOLIGNOL TRANSFERASE (PMT) do gene, resultando em uma maior frequência de resistente interunit obrigações e uma maior freqüência de G e S unidades de terminais com acesso de grupos fenólicos (Smith et al., 2015; Sibout et al., 2016). While in Arabidopsis the heterologous expression of PMT resulted in a reduced lignin amount accompanied by an increased saccharification efficiency (Sibout et al., 2016), não houve diminuição na quantidade de lignina no poplar e a eficiência de sacarificação não foi determinada (Smith et al., 2015).apesar de várias modificações da quantidade e composição de lignina terem sido demonstradas para melhorar o processamento da biomassa, estas modificações foram muitas vezes acompanhadas por uma penalização do Rendimento da biomassa (Leplé et al., 2007; Wadenbäck et al., 2008; Wagner et al., 2009; Voelker et al., 2010; Stout et al., 2014; Van Acker et al., 2014; Sykes et al., 2015; Zhou et al., 2018). A recent metastudy perturbed 21 lignin biosynthesis genes in P. trichocarpa, and comprehensively integrated the results of transcriptomic, proteomic, fluxomic, and phenomic data of 221 lines. Os autores concluíram que o crescimento da árvore não está associado com a quantidade de lignina, composição de subunidades ou ligações específicas (Wang et al., 2018), mas bastante correlacionado com a presença de navios Xylem Coleman et al.,2008a, b; Wagner et al., 2009; Voelker et al., 2010; Vargas et al., 2016; de Meester et al., 2018), a ativação de uma via de integridade Da Parede Celular (Bonawitz et al., 2014) e/ou a acumulação de inibidores químicos (Gallego-Giraldo et al., 2011; Muro-Villanueva et al., 2019).embora tenham sido envidados esforços substanciais para diminuir o teor de lignina através da redução da regulação dos genes biossintéticos da lignina, foram escassos os estudos sobre a regulação da via da lignina e a sobreprodução da lignina. Com efeito, os relatórios sobre a sobreexpressão do F5H revelam uma diminuição ou mesmo uma alteração do teor de lignina (Huntley et al., 2003; Li L. et al., 2003; Stewart et al., 2009; Mansfield et al., 2012; Edmunds et al., 2017). A sobreexpressão de CAD e COMT resultou em silenciamento genético ao invés de regulação, ou nenhum efeito nos níveis de expressão foi detectado (Baucher et al., 1996; Lapierre et al., 1999; Jouanin et al., 2000; Leplé et al., 2007; Van Acker et al., 2014). A sobreexpressão do R2R3-MYB fatores de transcrição PtoMYB92, PtoMYB216, e PtoMYB74, resultou em mais xylem camadas mais espessas xylem paredes celulares, bem como ectópica deposição de lignina, e as plantas, acumulados 13-50% a mais de lignina (Tian P. et al.= = Ligações externas = = , 2015; Li et al., 2018). O MYB sobreexpressão linhas constitutivamente upregulated a biossíntese de lignina caminho genes, e enquanto plantas overexpressing MYB92 e MYB74 tinha uma biomassa de penalidade, a sobreexpressão de MYB216 resultou em plantas com até 50% de lignina e não de desenvolvimento fenótipo. Como a lignina está sendo cada vez mais considerada um recurso importante para a produção sustentável de produtos químicos (Cao et al., 2018) a engenharia de plantas que produzem lignina em excesso deve ser mais explorada.os exemplos discutidos acima mostram claramente que a engenharia da lignina através de baixo ou upregulação de genes da via fenilpropanóide – ou expressão de genes heterólogos – tem o potencial de aumentar a eficiência de processamento da biomassa lignocelulósica. Devido a razões práticas e regulatórias, a maioria dos estudos relatam dados obtidos a partir da análise de árvores cultivadas em uma estufa. No entanto, os experimentos com árvores cultivadas em uma estufa normalmente não levam em conta processos de desenvolvimento, tais como cessação do crescimento e dormência. Além disso, os experimentos com efeito de estufa não fornecem informações suficientes sobre a interação da planta projetada com fatores ambientais, como tipo de solo, vento e patógenos. Compreender essas interações é um passo importante na tradução dos resultados da pesquisa para aplicações comerciais. Na verdade, o conjunto de trabalho produzido por estudos para os quais foi concedida permissão para estabelecer ensaios de campo, destaca diferenças importantes no fenótipo entre as árvores de estufa e de campo cultivados. A tabela 1 resume os relatórios sobre os ensaios de campo realizados com árvores 4CL, CCoAOMT, CCR, COMT e CAD.confirmando o potencial da biomassa lignocelulósica modificada como substrato para aplicações, várias árvores cultivadas com engenharia de lignina mostraram melhorias no processamento da madeira. Os poplares baixaram para o CCoAOMT cultivado por 5 anos em um teste de campo em Pequim (China), mostrou um aumento da liberação de glicose e xilose após a sacarificação (Wang et al., 2012). Os Poplars baixaram a regulamentação para a CCR e cresceram num ensaio de campo em França, provando-se ser mais receptivos à polpagem química kraft (Leplé et al., 2007). Dois ensaios de campo adicionais realizados em França e na Bélgica com poplares desclassificados CCR resultaram em uma melhoria de até 160% na produção de etanol em ensaios simultâneos de sacarificação e fermentação (SSF); no entanto, as plantas tiveram até 50% de redução de biomassa (Van Acker et al., 2014). Ensaios de campo com poplar CAD-baixo-regulamentado também mostraram resultados promissores. Estas árvores apresentavam um pouco menos de lignina do que o tipo selvagem e mostraram-se mais receptivas à delinificação do kraft (Lapierre et al., 1999). Consistentemente, as mesmas linhas cultivadas em ensaios de campo em grande escala em França e no Reino Unido mostraram uma ligeira diminuição da quantidade de lignina e uma melhoria da polpa de kraft considerada comercialmente relevante, uma vez que as plantas necessitavam de 6% menos de álcali para obter uma desalinhagem semelhante à das árvores de tipo selvagem (Pilate et al., 2002).no entanto, relatórios contraditórios sobre o Rendimento da biomassa e a eficiência de processamento a jusante sugerem que estes parâmetros são altamente influenciados por factores ambientais. Enquanto um ensaio de campo realizado na China usando poplares 4CL com baixa regulamentação descobriu que, mesmo com uma diminuição de 28% no conteúdo de lignina em comparação com o tipo selvagem, as árvores tinham cerca de 8% de altura aumentada (Tian X. M. et al., 2013), consistente com estudos de efeito estufa (Hu et al., 1999), outros ensaios de campo descobriram que os poplares 4CL-downregulated tinham diminuído a biomassa e eram às vezes mesmo Anões (Voelker et al., 2010; Stout et al., 2014; Marchin et al., 2017). Os relatórios divergem igualmente no que se refere à eficiência do processamento a jusante da madeira derivada destes poplares cultivados em campo 4CL. Enquanto até 100% de aumento na recuperação de açúcar foi encontrado para árvores 4cl1-downregulated (35S-driven antisense 4CL construct) cultivada em um local de montanha nos Estados Unidos (Xiang et al., 2017), dados obtidos a partir de estudos de campo realizados em Oregon (Estados Unidos) descobriram que o antisenso silênciado com promotor Pt4CL1 4cl1 poplars não tinha nenhuma melhoria na eficiência de sacarificação em comparação com o tipo selvagem (Voelker et al., 2010). Da mesma forma, um estudo de longo prazo em Wenling (China), descobriu que 4CL-Poplars downregulated não mostrou uma melhoria significativa no rendimento de açúcar em comparação com o tipo selvagem (Wang et al., 2012). Em ambos os últimos casos, as árvores mostraram diminuições suaves na quantidade de lignina que não se traduziram em maior eficiência de processamento, potencialmente devido à maior concentração de extrativos que poderiam interferir com a atividade enzimática (Voelker et al., 2010).estudos de campo demonstraram que os fatores ambientais podem influenciar a lignificação e restaurar traços aos níveis de tipo selvagem, em comparação com os níveis alcançados quando as mesmas plantas foram cultivadas na estufa. Enquanto 4CL-desativado árvores tinham diminuição do conteúdo de lignina, quando cultivada em estufa condições, a análise da mesma 4CL antisense choupos, mas cresceu no campo, muitas vezes mostrado que o teor de lignina foi aumentado em relação aos gases de efeito-cultivadas árvores e às vezes até mesmo restaurados para o tipo selvagem níveis (Stout et al., 2014; Xiang et al., 2017). Da mesma forma, os níveis de lignina foram muito menos reduzidos em poplares deficientes em CCR quando eles foram cultivados no campo em comparação com quando eles foram cultivados na estufa (Van Acker et al., 2014). Pelo menos para os poplares deficientes em CCR, é possível que o nível mais elevado de lignificação das árvores cultivadas no campo se deva ao facto de as amostras de madeira terem sido colhidas durante o inverno. Quando o crescimento da árvore cessa no outono, as árvores ainda têm tempo de lignificar completamente as suas paredes celulares quando a árvore entra em dormência, em comparação com as árvores de estufa que desenvolvem novos xylem continuamente. A composição da lignina também tem mostrado diferir entre as árvores de lignina de estufa e de campo. 4CL-downregulated poplars grown in a field in North Carolina had lignin with a lower S / G than when the same lines were grown in the greenhouse (Stout et al., 2014).no seu conjunto, estes resultados mostram que os dados obtidos a partir de árvores cultivadas com efeito de estufa não podem ser facilmente extrapolados para árvores cultivadas em campo, apoiando a necessidade de experiências de campo em diferentes locais. Algumas linhas apresentaram uma penalidade de rendimento tornando-os menos interessantes para as aplicações, destacando a necessidade de uma melhor compreensão da Base molecular da penalidade de rendimento e o desenvolvimento de estratégias para superar este problema.
lignina tem demonstrado desempenhar um papel importante na resistência aos agentes patogénicos (Miedes et al., 2014; Zhao e Dixon, 2014), e desempenha um papel fundamental em permitir que a usina Transporte Água. Isto sugere que as modificações da lignina podem ter um impacto na tolerância ao stress das plantas. Embora seja necessária mais investigação para abordar totalmente esta possibilidade, a regulamentação de 4CL, COMT, e CAD em poplar não alterou drasticamente o desempenho alimentar de herbívoros alimentadores de folhas (Tiimonen et al., 2005; Brodeur-Campbell et al., 2006; Hjalten et al., 2013). O efeito da baixa regulamentação da COMT e da CAD No choupo nas interações plant-insects também foi avaliado nas árvores cultivadas no campo, e foi demonstrado que as árvores modificadas com lignina tinham uma incidência normal de insetos visitando e alimentando, bem como respostas normais a patógenos microbianos (Pilatos et al., 2002; Halpin et al., 2007). Estes resultados indicam que as árvores com lignina modificada não sofrem necessariamente mais do que as plantas de tipo selvagem de Pragas e doenças. No entanto, a caracterização do microbioma bacteriano da endosfera de madeira colhida a partir de poplares cultivados no campo, com a CCR-downregulated, demonstrou mudanças na comunidade bacteriana, presumivelmente devido à abundância alterada de metabolitos fenólicos específicos no xylem (Beckers et al., 2017).considerando o papel da lignina na função e estrutura xilem, as relações de água de alguns poplares modificados com lignina foram avaliadas. 4CL-poplars downregulated foram encontrados para ter condutividade hidráulica reduzida, potencialmente interferindo com o crescimento da planta (Marchin et al., 2017). Os experimentos de estresse hidráulico com poplares foram re-regulados para CCR, COMT ou CAD mostraram que estas plantas tinham uma menor resistência à cavitação, mantendo a condutividade hidráulica xylem normal e transporte de água (Awad et al., 2012). Estes resultados sugerem que o crescimento dos mutantes de lignina baixa pode ser influenciado pela disponibilidade de água. Quanto a qualquer novo híbrido obtido da reprodução clássica, são necessários testes de campo para avaliar o desempenho do campo e a tolerância de esforço das árvores de engenharia de lignina.o desempenho das plantas de engenharia de linhite parece ser altamente influenciado pelas condições ambientais. Não é claro, no entanto, se as diferenças observadas entre árvores cultivadas com efeito de estufa e árvores cultivadas em campo, ou entre árvores cultivadas em diferentes locais de campo, resultam de diferentes níveis de supressão genética ou da interação do traço de engenharia com o ambiente (GxE). Na verdade, a regulamentação instável é uma deficiência de técnicas de silenciamento genético que são baseadas em RNAi. Isto é testemunhado pela observação da variação no fenótipo vermelho xilem que é observado quando genes de biossíntese da lignina, tais como CAD, COMT, ou CCR, são sub-regulados. A coloração xilem vermelha muitas vezes não é uniforme em todo o xylem, mas sim aparece em manchas que refletem níveis variáveis de silenciamento de genes (Leplé et al., 2007; Voelker et al., 2010; Van Acker et al., 2014; Figura 2). Além disso, o uso de métodos de silenciamento de genes pode potencialmente resultar no silenciamento concomitante de membros da família de genes estreitamente relacionados – talvez em vários graus – turvando as interpretações e camuflando os efeitos da desregulação de genes individuais.

Figura 2. A regulação genética irregular por RNAi. Fenótipo xilem vermelho irregular observado em troncos de poplares deficientes em CCR (à direita) cultivados num ensaio de campo na Bélgica. O xylem vermelho indica áreas de regulamentação do CCR. A madeira de árvores selvagens é esbranquiçada (à esquerda).
estas questões podem agora ser facilmente superadas pelo uso de tecnologias de edição de genes baseadas em CRISPR que permitem mutações estáveis de perda de funções (knock-outs) em genes alvo específicos, permitindo a dissecação da função de genes individuais dentro das famílias. Por exemplo, o alvo de membros individuais da família de genes 4CL no poplar mostrou que o 4CL1 está relacionado com a lignificação, enquanto o 4CL2 está envolvido na produção de proantocianidina (Zhou et al., 2015). Além dos alelos knock-out, a edição de genes CRISPR também permite criar novos alelos que conferem redução parcial na atividade enzimática. Isto abre a possibilidade de afinar o nível de actividade enzimática residual e de contornar a penalidade de rendimento que é frequentemente observada quando a quantidade de lignina desce abaixo de um nível limiar. Outro caminho promissor para lignina de engenharia na floresta de árvores possível através CRISPR baseado no genoma de engenharia é a edição simultânea de vários genes (alelos empilhamento) para otimizar o processamento da biomassa eficiência, como exemplificado em Arabidopsis, onde o empilhamento de a transaldolase (tra) e comt mutações, o c4h e comt mutações, ou o 4cl e comt mutações resultou em aditivo e sinérgico melhorias na sacarificação de eficiência (de Vries et al., 2018). Na verdade, uma abordagem de sistemas em P. trichocarpa prevê que a regulamentação concomitante de PAL e CCoAOMT, ou PAL, C3’h E CCOAOMT irá melhorar substancialmente as propriedades da madeira e a libertação de açúcar (Wang et al., 2018).a utilização da edição do genoma baseada na CRISPR na melhoria das árvores para as indústrias da pasta de papel e do papel e da bio-refinaria, bem como para a produção de aromáticos de plataformas a partir da degradação hidrogenolítica da lignina, será mais valiosa quando esta tecnologia for estrategicamente combinada com outras técnicas de reprodução (Figura 3). De facto, já existem grandes variações na quantidade de lignina e na composição S/G em populações naturais de árvores florestais (Studer et al., 2011). Dado que ambos os traços afectam a libertação de glucose após a saccharificação (Yoo et al., 2018), explorando esta diversidade genética através da criação convencional, auxiliada por estudos de associações genómicas (GWAS) (Porth et al., 2013; Fahrenkrog et al., 2017; Liu et al., 2018), reprodução com alelos defeituosos raros (BRDA) (Vanholme et al., 2013a) ou seleção genômica (Yin et al., 2010; Muchero et al., 2015; Pawar et al., 2018; Xie et al., 2018), é uma estratégia valiosa para obter linhas que melhoraram a eficiência de processamento de madeira. Uma vez que as árvores de elite são obtidas por estes métodos de reprodução, a engenharia genética e a edição de genes específicos baseada em CRISPR é uma via muito promissora para melhorar ainda mais estes genótipos de elite sem quebrar a sua constituição genética e sem passar por longos ciclos de reprodução. Dada a queda iminente do clima, não temos mais tempo a perder na adoção dessas novas técnicas de reprodução em nossa corrida para a economia baseada na biologia.
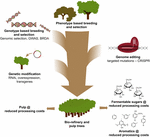
a Figura 3. Melhoramento genético de árvores florestais através de uma combinação de ferramentas de reprodução. Para acelerar a melhoria genética das árvores florestais para aplicações de celulose e biorrefinaria, as ferramentas clássicas e novas de reprodução precisam ser bem combinadas. A reprodução clássica envolve a selecção fenotípica de árvores para cruzamentos controlados, seguida de selecção fenotípica. Com o advento da Informação da sequência do genoma de muitas árvores florestais, novas estratégias como seleção genômica, estudos de associação de genoma amplo (GWAS) e criação com alelos raros defeituosos (BRDA) foram desenvolvidas para acelerar a captura e enriquecimento de polimorfismos de DNA associados com características benéficas. A edição baseada em CRISPR do genoma permite modificar o genoma de uma forma que imita polimorfismos naturais. A modificação genética envolve a integração estável de DNA estranho na árvore para superprodução (uma) enzimática(s) ou para o (s) gene(s) de downregulate (a). A combinação das técnicas clássicas e novas de reprodução é necessária para fornecer madeira de qualidade suficiente para a sociedade.
contribuições do autor
todos os autores listados fizeram uma contribuição substancial, direta e intelectual para a obra, e aprovaram-na para publicação.
financiamento
reconhecemos o financiamento parcial do BIOLEUM do projecto IWT-SBO (subvenção n. º 130039), e pela SBO-FISH através do projecto ARBOREF. A AC recebeu financiamento da FWO e do Programa de investigação e inovação Horizonte 2020 da União Europeia ao abrigo da Convenção de subvenção Marie Skłodowska-Curie n. o 665501. A LdV foi financiada pelo Instituto para a promoção da inovação através da Ciência e da tecnologia na Flandres (IWT-Vlaanderen) para uma bolsa de estudo pré-Doutoral.
Declaração de conflito de interesses
os autores declaram que a investigação foi realizada na ausência de quaisquer relações comerciais ou financeiras que possam ser interpretadas como um potencial conflito de interesses.
agradecimentos
Agradecemos a Annick Bleys por preparar este manuscrito para a submissão.
Boerjan, W., Ralph, J., and Baucher, M. (2003). Biossíntese da lignina. Annu. Rev. Plant Biol. 54, 519–546.
PubMed Resumo | Google Scholar
Lapierre, C., Pollet, B., Petit-Conil, M., Toval, G., Romero, J., Pilatos, G., et al. (1999). As alterações estruturais da lignina em poplares transgénicos com álcool desidrogenase de canela depressivo ou actividade o-metiltransferase de ácido caffeico têm um impacto oposto na eficiência da polpagem industrial de kraft. Plant Physiol. 119, 153–163.
PubMed Abstract/Google Scholar
Ralph, J. (2006). “O que faz um bom substituto do monolignol?, “in the Science and Lore of the Plant Cell Wall-Biosynthesis, Structure and Function, ed. T. Hayashi (Boca Raton, FL: BrownWalker Press).
Google Scholar