Inledning
fossila resurser är den viktigaste råvaran för energi och organiska föreningar, och deras användning resulterar i utsläpp av växthusgaser i samband med klimatförändringar. Den kommande klimatkraschen kräver en brådskande övergång från en fossilbaserad till en biobaserad ekonomi där lignocellulosisk biomassa snarare än olja används för produktion av bränslen, kemikalier och material. Trä är en viktig källa till lignocellulosisk biomassa; den består huvudsakligen av sekundära förtjockade cellväggar rik på cellulosa, hemicelluloser och lignin. Alla tre polymererna kan värdesättas i den biobaserade ekonomin. Cellulosa är en källa för massa-och pappersindustrin, och både cellulosa och hemicellulosa kan depolymeriseras till deras monosackarider för jäsning till t.ex. bioetanol, mjölksyra och tvättmedel (Vanholme et al., 2013b). Eftersom lignin negativt påverkar effektiviteten av träbearbetning mot dessa applikationer kan träd konstrueras för att ackumulera mindre lignin, för att bli mer mottagliga för produktion av papper och fermenterbara sockerarter. Å andra sidan betraktas lignin alltmer som en värdefull komponent i den biobaserade ekonomin. Med tanke på att lignin är den största förnybara aromatiska källan på jorden, kan den ekonomiska livskraften hos ett bioraffinaderi ökas avsevärt om lignin också valoriseras och används som en resurs för produktion av kemikalier (Holladay et al., 2007; Tuck et al., 2012; Davis et al., 2013; Ragauskas et al., 2014; Li C. et al., 2015; Van Den Bosch et al., 2015; Rinaldi et al., 2016; Upton och Kasko, 2016; Schutyser et al., 2018).
ligninpolymeren består av monolignoler som produceras av fenylpropanoid-och monolignolbiosyntetiska vägar, genom en serie enzymatiska reaktioner som börjar med deaminering av fenylalanin (Figur 1). Monolignolerna syntetiseras i cytoplasman och translokeras till apoplasten, där de dehydrogeneras till monolignolradikaler genom verkan av lakfall och peroxidaser (Berthet et al., 2011; Zhao et al., 2013). Dessa monolignolradikaler parar sedan med varandra på ett kombinatoriskt sätt och genererar en rad kemiska bindningar såsom aryl-eterbindningen (exportorienterad-o-4), resinolbindningen (exportorienterad-exportorienterad) och fenylkoumaranbindningen (exportorienterad-5) (Boerjan et al., 2003; Ralph et al., 2004; Vanholme et al., 2010). De vanligaste monolignolerna är hydroxycinnamylalkoholerna p-kumaryl, coniferyl och sinapylalkoholer, som genererar h -, G-och S-enheterna vid deras införlivande i ligninpolymeren (Bonawitz och Chapple, 2010; Ralph et al., 2019; Vanholme et al., 2019). Det relativa bidraget från ligninbyggnadsblocken varierar mellan taxa, utvecklingsstadium, vävnad och celltyp och till och med cellväggskikt; lignin från barrved (gymnospermer) består nästan helt av G-enheter med en mindre bråkdel av H-enheter, medan lignin från lövträ (angiospermer) har s-enheter utöver G-enheter och spår av H-enheter (Boerjan et al., 2003; Vanholme et al., 2010, 2019). Förutom dessa traditionella monolignoler kan en mängd andra p-hydroxylerade aromatiska molekyler införlivas i ligninpolymeren till olika nivåer (Vanholme et al., 2019).
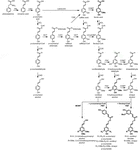
Figure 1. Lignin biosynthetic pathway. Alternative monomers and heterologously expressed enzymes are shown in bold. PAL, phenylalanine ammonia-lyase; C4H, cinnamate 4-hydroxylase; 4CL, 4-coumarate:CoA ligase; HCT, p-Hydroxycinnamoyl-CoA:quinate/shikimate-p-hydroxycinnamoyltransferase; C3’H, p-coumaroyl quinate/shikimate 3’-hydroxylase; CSE, caffeoyl shikimate esterase; CCoAOMT, caffeoyl-CoA O-methyltransferase; CCR, cinnamoyl-CoA reductase; F5H, ferulate 5-hydroxylase/CAld5H, coniferaldehyde 5-hydroxylase; COMT, koffeinsyrao-metyltransferas; CAD, kanelalkoholdehydrogenas; FMT, feruloyl-CoA monolignoltransferas; PMT, p-kumaroyl-CoA monolignoltransferas.
med tanke på att lignin är en viktig recalcitrancefaktor i trädelignifieringsprocesser har stora forskningsinsatser ägnats åt att avslöja ligninbiosyntetiska vägen och att studera effekterna av störningar av ligninbiosyntesgenerna på ligninmängd och sammansättning och på träförädlingseffektivitet. Medan modifieringar i gener som sträcker sig från de kodande transkriptionsfaktorerna upp till de kodande oxidativa enzymerna har resulterat i förändrat lignininnehåll, sammansättning eller avsättning (Eriksson et al., 2000; Li Y. H. et al., 2003; Liang et al., 2008; Lu et al., 2013; Lin et al., 2016; Xu et al., 2017; Yang et al., 2017; Obudulu et al., 2018) kommer denna översyn att fokusera på resultaten som erhållits genom att konstruera ligninbiosyntetiska gener.
Engineering Ligninvägen
tabell 1 ger en översikt över de olika studierna på nedreglerade eller muterade ligninbiosyntetiska gener i poppel, tall, eukalyptus och björk, med de resulterande effekterna på träbearbetningseffektivitet, när de bestäms. Att minska aktiviteten för varje steg i ligninbiosyntesvägen, från PAL upp till CAD, kan leda till en minskning av ligninhalten (Tabell 1). Flera parametrar påverkar graden av ligninreduktion, såsom målgenen och graden av nedreglering av enzymaktiviteten, vilket i sin tur beror på effektiviteten hos den använda ljuddämpningskonstruktionen, genfamiljens storlek och redundans inom genfamiljen. Generellt resulterar nedregleringen av stegen från C4H upp till CCR i en mer dramatisk minskning av ligninmängden (Hu et al., 1999; Meyermans et al., 2000; Zhong et al., 2000; Li L. et al., 2003; Jia et al., 2004; Lu et al., 2004; LEPL 6 et al., 2007; Coleman et al., 2008a, b; Bjurhager et al., 2010; Mansfield et al., 2012; Ralph et al., 2012; Min et al., 2014; Van Acker et al., 2014; Zhou et al., 2015, 2018; Saleme et al., 2017; Xiang et al., 2017) än nedreglering av F5H, COMT och CAD (Van Doorsselaere et al., 1995; Baucher et al., 1996; Lapierre et al., 1999; Jouanin et al., 2000; Van Acker et al., 2014; Wang et al., 2018). Ligninreduktion kan associeras med en ökning av S/G, såsom i C3 ’ h- (Coleman et al., 2008a; Ralph et al., 2012) och CCoAOMT-nedreglerade träd (Meyermans et al., 2000), eller en minskning av s/G – förhållandet, såsom i CSE – (Saleme et al., 2017), och COMT-nedreglerade träd (Van Doorsselaere et al., 1995; Lapierre et al., 1999; Jouanin et al., 2000). Intressant, låg-lignin 4CL-downregulated poplars befanns ha en ökning av S / g (Min et al., 2014; Xiang et al., 2017), en minskning av S/G (Voelker et al., 2010; Zhou et al., 2015), eller förhållanden jämförbara med vildtyp (Hu et al., 1999; Li L. et al., 2003). Denna varians kan inte associeras med promotorn eller metoden som används för nedreglering, vilket tyder på att skillnader i graden av tystnad, tillväxtförhållanden eller utvecklingstillstånd påverkar detta drag. Å andra sidan har de starkaste effekterna på H/G/S ligninsammansättning observerats för träd nedreglerade i C3 ’ H och HCT, som deponerar lignin med stora ökningar av H-enhetens innehåll (Coleman et al., 2008a; Ralph et al., 2012; Vanholme et al., 2013a), medan träd som överuttrycker F5H producerar lignin starkt berikat i S-enheter (Franke et al., 2000; Li L. et al., 2003; Stewart et al., 2009), och träd som är nedreglerade i COMT har dramatiskt minskat s-enhetens biosyntes (Van Doorsselaere et al., 1995; Lapierre et al., 1999).

tabell 1. Översikt över skogsträd med modifierat uttryck av ligninbiosyntesgener.
biomassans bearbetningseffektivitet kan också modifieras genom ökad inkorporering av molekyler som i allmänhet representerar mindre komponenter i lignin av vilda växter. Införlivandet av ferulsyra i CCR-bristfälliga träd resulterar i bildandet av acetalbindningar i ligninpolymeren, som lätt klyvs i sura biomassapreparat (LEPL Ukrainian et al., 2007; Ralph et al., 2008; Van Acker et al., 2014). Faktum är att nivåerna av ferulsyra i lignin korrelerade positivt med en högre sackarifieringseffektivitet (Van Acker et al., 2014). Införlivandet av 5-hydroxikoniferylalkohol och 5-hydroxikoniferaldehyd i lignin av COMT-bristande Poplar (Van Doorsselaere et al., 1995; Lapierre et al., 1999; Jouanin et al., 2000; Morreel et al., 2004; Lu et al., 2010) ger upphov till bensodioxanbindningar, vilket potentiellt förhindrar kovalenta kopplingar mellan lignin och polysackaridhydroxylgrupperna (Weng et al., 2010; Vanholme et al., 2012; Nishimura et al., 2018). Å andra sidan resulterar COMT-brist också i ett mer kondenserat lignin på grund av de relativt högre halterna av de kondenserade obligationerna mellan export och export och 5 och de lägre nivåerna av obligationerna mellan export och export, När s-enhetsfrekvensen sjunker. Kemisk massa av trä härrörande från Poplar kraftigt nedreglerad för COMT resulterade i ett högre massavkastning, motvikt av kvarvarande ligninhalt i massan. Dessa träd hade en lägre lignin och en högre cellulosahalt (Jouanin et al., 2000). Å andra sidan hade Poplar som var blygsamt nedreglerade för COMT en stor minskning av massavkastningen, förmodligen för att lignininnehållet hade förblivit normalt medan ligninet hade en högre frekvens av kondenserade bindningar som negativt påverkade ligninextraktionen (Lapierre et al., 1999; Pilatus et al., 2002). Införlivandet av kaneldehyder i ligninpolymeren i CAD-bristfälliga träd resulterar i kortare ligninpolymerkedjor, därav en högre andel fria fenoliska ändgrupper som ökar lösligheten hos polymeren i alkali. Införlivandet av kaneldehyder i ligninpolymeren minskar förmodligen också den kovalenta interaktionen mellan den alifatiska kedjan och hemicellulosa, vilket återigen gör ligninet mer lösligt. Dessutom, på grund av det utvidgade konjugerade systemet som genereras när en kanelaldehyd Bisexuell-O-4 parar med en annan monomer, blir den aromatiska eterbindningen av den inkorporerade kanelaldehyden mer mottaglig för alkalisk klyvning (Lapierre et al., 1989; Van Acker et al., 2017).
Ligninpolymerisation är en kombinatorisk radikal kopplingsprocess, vilket gör att ett brett spektrum av fenolföreningar naturligt kan införlivas i ligninpolymeren (Boerjan et al., 2003; Vanholme et al., 2019). Forskare har försökt att skräddarsy ligninmängden och kompositionen för att förbättra biomassabearbetning genom uttryck av heterologa gener, som syftar till biosyntes och införlivande av olika kompatibla fenolföreningar som alternativa monolignoler i ligninpolymeren (Ralph, 2006; Vanholme et al., 2012; Mottiar et al., 2016; Mahon och Mansfield, 2018). Ett exempel är införandet av gener som kodar för enzymer som behövs för S-enhetens biosyntes i Tall; det samtidiga uttrycket av F5H, COMT och CAD introducerade framgångsrikt s-enheter i Pinus radiata (Wagner et al., 2015; Edmunds et al., 2017). Införandet av genen som kodar för ett monolignol 4-O-metyltransferas (MOMT4) i poppel leder till bildandet av 4-o-metylerade koniferyl-och sinapylalkoholer, som inte kan införlivas i den växande ligninpolymeren eftersom de saknar den aromatiska hydroxylgruppen. Detta leder till ett stopp i ligninpolymerisation och resulterar i träd med lägre ligninhalt och högre sackarifieringseffektivitet (Bhuiya och Liu, 2010; Cai et al., 2016). Poplars har också konstruerats för att innehålla esterbindningar i ligninpolymerens ryggrad. Coniferylferulatestrar infördes i polymeren via uttryck av en FERULOYL-CoA:MONOLIGNOLTRANSFERAS (FMT) gen härledd från Angelica sinensis (Wilkerson et al., 2014), vilket leder till en förbättrad sackarifieringseffektivitet under olika förbehandlingsförhållanden (Wilkerson et al., 2014; Kim et al., 2017; Bhalla et al., 2018), och en förbättrad kraftmasseffektivitet jämfört med vildtyp (Zhou et al., 2017). Monolignol p-kumaratestrar har också konstruerats i poppel, via uttryck av en ris p-KUMAROYL-CoA:MONOLIGNOLTRANSFERAS (PMT) gen, vilket resulterar i en högre frekvens av resistenta interenhetsbindningar och en högre frekvens av G-och s-terminalenheter med fria fenolgrupper (Smith et al., 2015; Sibout et al., 2016). Medan i Arabidopsis resulterade det heterologa uttrycket av PMT i en reducerad lignin mängd åtföljd av en ökad sackarifieringseffektivitet (Sibout et al., 2016) fanns det ingen minskning av ligninmängden i poppel och sackarifieringseffektiviteten bestämdes inte (Smith et al., 2015).
medan flera modifieringar av ligninmängden och sammansättningen visade sig ge förbättringar i biomassaförädling, åtföljdes dessa modifieringar ofta av en straff för biomassaavkastning (LEPL Jacobsen et al., 2007; Wadenb Jacobck et al., 2008; Wagner et al., 2009; Voelker et al., 2010; Stout et al., 2014; Van Acker et al., 2014; Sykes et al., 2015; Zhou et al., 2018). En ny metastudie störde 21 ligninbiosyntesgener I P. trichocarpa, och omfattande integrerade resultaten av transkriptomiska, proteomiska, fluxomiska och fenomenala data om 221 linjer. Författarna drog slutsatsen att trädtillväxt inte är associerad med ligninmängd, subenhetskomposition eller specifika kopplingar (Wang et al., 2018), men snarare korrelerad med närvaron av kollapsade xylemfartyg (Coleman et al., 2008a, b; Wagner et al., 2009; Voelker et al., 2010; Vargas et al., 2016; de Meester et al., 2018), aktiveringen av en cellväggintegritetsväg (Bonawitz et al., 2014) och / eller ackumulering av kemiska hämmare (Gallego-Giraldo et al., 2011; Muro-Villanueva et al., 2019).medan betydande ansträngningar har gjorts för att minska ligninhalten genom nedreglering av ligninbiosyntetiska gener, har studier om uppreglering av ligninvägen och överproduktionen av lignin varit knappa. Faktum är att rapporter om överuttryck av F5H visar en oförändrad eller till och med en minskning av ligninhalten (Huntley et al., 2003; Li L. et al., 2003; Stewart et al., 2009; Mansfield et al., 2012; Edmunds et al., 2017). Överuttrycket av CAD och COMT har resulterat i gen-tystnad snarare än uppreglering, eller ingen effekt på uttrycksnivåer detekterades (Baucher et al., 1996; Lapierre et al., 1999; Jouanin et al., 2000; LEPL Bisexuell et al., 2007; Van Acker et al., 2014). Överuttrycket av r2r3-MYB-transkriptionsfaktorerna PtoMYB92, PtoMYB216 och PtoMYB74 resulterade alla i ytterligare xylemskikt, tjockare xylemcellväggar samt ektopisk ligninavsättning och växterna ackumulerade 13-50% mer lignin (Tian Q. et al., 2013; Li C. F. et al., 2015; Li et al., 2018). MYB-överuttryckslinjerna uppreglerade konstitutivt ligninbiosyntesväggenerna, och medan växter som överuttryckte MYB92 och MYB74 hade en biomassastraff, resulterade ÖVERUTTRYCKET av MYB216 i växter med upp till 50% mer lignin och ingen utvecklingsfenotyp. Eftersom lignin alltmer betraktas som en viktig resurs för hållbar produktion av kemikalier (Cao et al., 2018) konstruktionen av växter som överproducerar lignin bör undersökas ytterligare.
fältförsök
exemplen som diskuterats ovan visar tydligt att ligninteknik via ned – eller uppreglering av fenylpropanoidvägsgener – eller uttryck av heterologa gener-har potential att öka bearbetningseffektiviteten hos lignocellulosisk biomassa. På grund av praktiska och reglerande skäl rapporterar de flesta studier om data som erhållits från analysen av träd som odlas i ett växthus. Experiment med träd som odlas i ett växthus tar emellertid vanligtvis inte hänsyn till utvecklingsprocesser som tillväxtstopp och viloläge. Dessutom ger växthusexperiment inte tillräcklig inblick i interaktionen mellan den konstruerade växten och miljöfaktorer som jordtyp, vind och patogener. Att förstå dessa interaktioner är ett viktigt steg i översättningen av forskningsresultat mot kommersiella tillämpningar. Verkligen, kroppen av arbete som produceras av studier för vilka tillstånd att etablera fältförsök beviljades, belyser viktiga skillnader i fenotyp mellan växthus-och fält odlade träd. Tabell 1 sammanfattar rapporterna om fältförsök utförda med 4CL, CCoAOMT, CCR, COMT och CAD nedreglerade träd.
bekräftar potentialen hos modifierad lignocellulosisk biomassa som substrat för applikationer, flera lignin-konstruerade fältförsöksodlade träd visade förbättringar i träförädling. Poplars nedreglerade för CCoAOMT odlade i 5 år i ett fältförsök i Peking (Kina), visade en ökad glukos och xylos frisättning vid sackarifiering (Wang et al., 2012). Poplars nedreglerade för CCR och odlas i ett fältförsök i Frankrike, visade sig vara mer mottagliga för kemisk kraft massa (LEPL Ukrainian et al., 2007). Två ytterligare fältförsök som genomfördes i Frankrike och Belgien med CCR-nedreglerade Poplar resulterade i upp till 160% förbättring av etanolproduktionen vid samtidig sackarifiering och jäsning (SSF) analyser; växterna hade dock upp till 50% minskning av biomassa (Van Acker et al., 2014). Fältförsök med CAD-nedreglerad poppel visade också lovande resultat. Dessa träd visade något mindre lignin än vildtyp och visade sig vara mer mottagliga för kraft delignification (Lapierre et al., 1999). Konsekvent visade samma linjer som odlades i större fältförsök i Frankrike och Storbritannien en mild minskning av ligninmängden och en förbättring av kraftmassa som ansågs kommersiellt relevant, eftersom växterna behövde 6% mindre alkali för att uppnå en delignifiering som liknar den för vildtypsträd (Pilate et al., 2002).
motstridiga rapporter om både biomassaavkastning och bearbetningseffektivitet i efterföljande led tyder dock på att dessa parametrar påverkas starkt av miljöfaktorer. Medan en fältförsök som genomfördes i Kina med 4CL nedreglerade poplars fann att även med en 28% minskning av ligninhalten jämfört med vildtyp hade träden cirka 8% ökad höjd (Tian XM et al., 2013), i överensstämmelse med växthusstudier (Hu et al., 1999) fann andra fältförsök att 4CL-nedreglerade Poplar hade minskat biomassa och ibland till och med dvärgades (Voelker et al., 2010; Stout et al., 2014; Marchin et al., 2017). Rapporter skiljer sig också om nedströms bearbetningseffektivitet av trä som härrör från dessa 4CL-nedreglerade fältodlade Poplar. Medan upp till 100% ökning av sockeråtervinning hittades för 4cl1-nedreglerade träd (35S-driven antisense 4CL-konstruktion) odlade på en bergsplats i USA (Xiang et al., 2017), data erhållna från fältstudier utförda i Oregon (USA) fann att pt4cl1 promotordrivna antisense tystade 4cl1 poplars inte hade någon förbättring av sackarifieringseffektiviteten jämfört med vildtyp (Voelker et al., 2010). På samma sätt fann en långsiktig studie i Wenling (Kina) att 4CL-nedreglerade Poplar inte visade en signifikant förbättring av sockerutbytet jämfört med vildtyp (Wang et al., 2012). I båda senare fallen visade träden milda minskningar i ligninmängd som inte översattes till högre bearbetningseffektivitet, potentiellt på grund av den högre koncentrationen av extraktionsmedel som kan störa enzymatisk aktivitet (Voelker et al., 2010).
fältstudier har visat att miljöfaktorer kan påverka lignifiering och återställa egenskaper till vildtypsnivåer jämfört med de nivåer som uppnåddes när samma växter odlades i växthuset. Medan 4CL-nedreglerade träd hade minskat ligninhalten när de odlades under växthusförhållanden, analys av samma 4CL antisense poplars, men odlade i fältet, har ofta visat att ligninhalten ökades jämfört med de växthusodlade träden och ibland till och med återställdes till vildtypsnivåer (Stout et al., 2014; Xiang et al., 2017). På samma sätt reducerades ligninhalterna mycket mindre i CCR-bristfälliga Poplar när de odlades i fältet jämfört med när de odlades i växthuset (Van Acker et al., 2014). Åtminstone för de CCR-bristfälliga poplarna är det möjligt att den högre lignifieringsnivån för fältodlade träd beror på att träproverna togs under vintern. När trädtillväxten upphör på hösten har träden fortfarande tid att helt lignifiera sina cellväggar när trädet går in i viloläge, jämfört med växthusodlade träd som kontinuerligt utvecklar nytt xylem. Lignin sammansättning har också visat sig skilja mellan växthus-och fält odlade låg lignin träd. 4CL-nedreglerade Poplar odlade i ett fält i North Carolina hade lignin med lägre S/G än när samma linjer odlades i växthuset (Stout et al., 2014).sammantaget visar dessa resultat att data som erhållits från växthusodlade träd inte lätt kan extrapoleras till fältodlade träd, vilket ligger till grund för behovet av fältförsök på olika platser. Vissa linjer presenterade en avkastningsstraff som gjorde dem mindre intressanta för applikationer, vilket belyser behovet av en bättre förståelse för den molekylära grunden för avkastningsstraffet och utvecklingen av strategier för att övervinna detta problem.
Lignin har visat sig spela en viktig roll i patogenresistens (Miedes et al., 2014; Zhao och Dixon, 2014), och det spelar en central roll för att låta anläggningen transportera vatten. Detta tyder på att lignin modifieringar kan ha en inverkan på växtstress tolerans. Medan ytterligare undersökning behövs för att fullt ut ta itu med denna möjlighet förändrade nedregleringen av 4cl, COMT och CAD i poppel inte dramatiskt utfodringsprestanda för bladmatande växtätare (Tiimonen et al., 2005; Brodeur-Campbell et al., 2006; Hjalten et al., 2013). Effekten av nedreglering av COMT och CAD i poppel på växt-insektsinteraktioner har också bedömts på fältvuxna träd, och det visades att de lignin-modifierade träden hade normal förekomst av besökande och utfodringsinsekter, liksom normala svar på mikrobiella patogener (Pilate et al., 2002; Halpin et al., 2007). Dessa resultat indikerar att träd med modifierat lignin inte nödvändigtvis lider mer än vilda växter från skadedjur och sjukdomar. Icke desto mindre visade profilering av endosfärens bakteriella mikrobiom av trä skördat från fältodlade, CCR-nedreglerade poplars förändringar i bakteriesamhället, förmodligen på grund av det förändrade överflödet av särskilda fenolmetaboliter i xylem (Beckers et al., 2017).
Med tanke på lignins roll i xylemfunktion och struktur har vattenförhållandena hos några få låg lignin-modifierade Poplar utvärderats. 4CL-nedreglerade Poplar visade sig ha minskad hydraulisk ledningsförmåga, potentiellt störande växttillväxt (Marchin et al., 2017). Hydrauliska spänningsexperiment med poplars nedreglerade för CCR, COMT eller CAD visade att dessa växter hade ett lägre motstånd mot kavitation, samtidigt som normal xylem hydraulisk ledningsförmåga och vattentransport bibehölls (Awad et al., 2012). Dessa resultat tyder på att tillväxten av låg ligninmutanter kan påverkas av vattentillgänglighet. Som för varje ny hybrid erhållen från klassisk avel behövs fälttester för att utvärdera fältprestanda och stresstolerans hos lignin-konstruerade träd.
utsikter för Ligninteknik i skogsträd
prestandan hos ligninkonstruerade växter verkar påverkas starkt av miljöförhållandena. Det är emellertid oklart huruvida skillnaderna som observerats mellan växthusodlade och fältodlade träd eller mellan träd som odlas på olika fältplatser beror på olika nivåer av genundertryckning eller från interaktion mellan den konstruerade egenskapen med miljön (GxE). Faktum är att instabil nedreglering är en brist på gendämpningstekniker som är baserade på RNAi. Detta bevittnas genom att observera variation i den röda xylemfenotypen som observeras när specifika ligninbiosyntesgener, såsom CAD, COMT eller CCR, nedregleras. Den röda xylemfärgningen är ofta inte enhetlig i hela xylemen, utan förekommer snarare i fläckar som återspeglar variabla nivåer av gentystning (LEPL Bisexuell et al., 2007; Voelker et al., 2010; Van Acker et al., 2014; Figur 2). Dessutom kan användningen av genljuddämpningsmetoder potentiellt resultera i samtidig tystnad av närbesläktade genfamiljmedlemmar – kanske i olika grad – grumla tolkningarna och kamouflera effekterna av nedreglering av enskilda gener.

Figur 2. Patchy gen nedreglering av RNAi. Fläckig röd xylemfenotyp observerad på stammar av CCR-bristfälliga Poplar (höger) odlade i ett fältförsök i Belgien. Den röda xylemen indikerar områden med CCR-nedreglering. Trä från vilda träd är vitaktig (vänster).
dessa problem kan nu enkelt övervinnas genom användning av CRISPR-baserad genredigeringsteknik som möjliggör stabila funktionsförlustmutationer (knock-outs) i specifika målgener, vilket möjliggör dissektion av funktionen hos enskilda gener inom familjer. Till exempel visade inriktningen av enskilda 4CL-genfamiljmedlemmar i poppel att 4CL1 är relaterad till lignifiering, medan 4CL2 är involverad i proanthocyanidinproduktion (Zhou et al., 2015). Förutom knock-out alleler tillåter CRISPR-baserad genredigering också att skapa nya alleler som ger partiell minskning av enzymaktivitet. Detta öppnar möjligheten att finjustera nivån på kvarvarande enzymaktivitet och att kringgå avkastningsstraffet som ofta observeras när ligninmängden sjunker under en tröskelnivå. En annan lovande aveny för ligninteknik i skogsträd möjliggjort genom CRISPR-baserad genomteknik är samtidig redigering av flera gener (allelstapling) för att optimera biomassans bearbetningseffektivitet, vilket exemplifieras i Arabidopsis där stapling av transaldolas (tra) och comt-mutationer, C4H-och comt-mutationerna eller 4CL-och comt-mutationerna resulterade i additiva och synergistiska förbättringar i sackarifieringseffektivitet (de Vries et al., 2018). Verkligen, ett system tillvägagångssätt i P. trichocarpa förutspår att samtidig nedreglering av PAL och CCoAOMT, eller PAL, C3 ’ H och CCOAOMT avsevärt kommer att förbättra träegenskaper och sockerfrisättning (Wang et al., 2018).
användningen av CRISPR – baserad genomredigering i trädförbättring för massa – och pappers-och bioraffinaderiindustrin, liksom för produktion av plattformsaromater från hydrogenolytisk nedbrytning av lignin, kommer att vara mest värdefull när denna teknik strategiskt kombineras med andra avelstekniker (Figur 3). Faktum är att stor variation i ligninmängd och s/G-Sammansättning redan finns i naturliga populationer av skogsträd (Studer et al., 2011). Med tanke på att båda egenskaperna påverkar glukosfrisättningen vid sackarisering (Yoo et al., 2018), utnyttja denna genetiska mångfald genom konventionell avel, med hjälp av Genome Wide Association Studies (GWAS) (Porth et al., 2013; Fahrenkrog et al., 2017; Liu et al., 2018), avel med sällsynta defekta alleler (BRDA) (Vanholme et al., 2013a) eller genomiskt urval (Yin et al., 2010; Muchero et al., 2015; Pawar et al., 2018; Xie et al., 2018), är en värdefull strategi för att få linjer som har förbättrat träbearbetningseffektiviteten. När elitträd erhålls genom dessa avelsmetoder är genteknik och CRISPR-baserad genredigering av specifika gener en mycket lovande väg att ytterligare förbättra dessa elitgenotyper utan att bryta upp sin genetiska konstitution och utan att gå igenom långa avelscykler. Med tanke på den överhängande klimatkraschen har vi inte mer tid att förlora när vi antar dessa nya avelstekniker i vår ras till den biobaserade ekonomin.
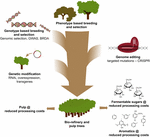
Figur 3. Genetisk förbättring av skogsträd genom en kombination av avelsverktyg. För att påskynda den genetiska förbättringen av skogsträd för massa-och bioraffinaderiapplikationer måste klassiska och nya avelsverktyg kombineras smart. Klassisk avel innebär fenotypiskt urval av träd för kontrollerade kors, följt av fenotypiskt urval. Med tillkomsten av genomsekvensinformation av många skogsträd har nya strategier som genomiskt urval, Genome Wide Association Studies (GWAS) och avel med sällsynta defekta alleler (BRDA) utvecklats för att påskynda infångningen och berikningen av DNA-polymorfier associerade med fördelaktiga egenskaper. CRISPR-baserad genomredigering gör det möjligt att modifiera genomet på ett sätt som efterliknar naturliga polymorfier. Genetisk modifiering innebär stabil integration av främmande DNA i trädet för att överproducera (an) enzym(er) eller nedreglera (a) gen(er). Att kombinera de klassiska och nya Avelsteknikerna behövs för att ge tillräckligt högkvalitativt trä för samhället.
Författarbidrag
alla listade författare har gjort ett väsentligt, direkt och intellektuellt bidrag till arbetet och godkänt det för publicering.
finansiering
Vi erkänner delfinansiering från IWT-SBO-projektet BIOLEUM (bidrag nr 130039) och av SBO-FISH genom ARBOREF-projektet. AC har erhållit finansiering från FWO och EU: s forsknings-och innovationsprogram Horizon 2020 inom ramen för Marie Sk Portuguodowska-Curie bidragsavtal nr 665501. LdV finansierades av Institutet för främjande av Innovation genom vetenskap och teknik i Flandern (IWT-Vlaanderen) för ett predoctoralt stipendium.
intressekonflikt uttalande
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
bekräftelser
Vi tackar Annick Bleys för att förbereda detta manuskript för inlämning.
Boerjan, W., Ralph, J. och Baucher, M. (2003). Lignin biosyntes. Annu. Rev. Växt Biol. 54, 519–546.
PubMed Abstract / Google Scholar
Lapierre, C., Pollet, B., Petit-Conil, M., Toval, G., Romero, J., Pilatus, G., et al. (1999). Strukturella förändringar av ligniner i transgena Poplar med deprimerad kanelalkoholdehydrogenas eller koffeinsyra-o-metyltransferasaktivitet har en motsatt inverkan på effektiviteten hos industriell kraftmassa. Växt Physiol. 119, 153–163.
PubMed Abstract / Google Scholar
Ralph, J. (2006). ”Vad gör en bra monolignol substitut?, ”i vetenskapen och Lore av Växtcellväggen – biosyntes, struktur och funktion, Red. T. Hayashi (Boca Raton, FL: BrownWalker Press).
Google Scholar