introduktion
framgångsrik interaktion med vår sensoriska miljö kräver en invecklad balans mellan två attentional selection-mekanismer-den av top-down och bottom-up. Rubrik över till producera gången i din lokala stormarknad med målet att plocka upp några nödvändiga ingredienser för mango sallad, engagerar utplacering av frivilliga målstyrda, eller top-down, attentional system så att du aktivt söka efter alla nödvändiga ingredienser bland de många producera val. Men om du hör en ringsignal på en mobiltelefon kommer det sannolikt att fånga din uppmärksamhet och avbryta din sökning. Ett sådant avbrott sker i en bottom-up, eller stimulusdriven, mode där en ren salience av stimulus, det faktum att ringen skiljer sig från andra ljud i din miljö, anser det värt val. Det beskrivna scenariot understryker vikten av målinriktat och stimulansdrivet urval för beteende och pekar på en fin balans som måste existera mellan de två uppmärksamhetssystemen för att förhindra ”tunnelsyn” å ena sidan och fullständig oförmåga att fokusera på den andra.
Top-Down och Bottom-Up Val: Beteende
flera decennier av beteendeforskning har ägnats åt att visa att fördelningen av uppmärksamhet kan styras av observatörens avsikter såväl som av den fysiska stimulansens framträdande. Mycket av beteendebevis för top-down och bottom-up attentional allocation har granskats i stor utsträckning någon annanstans (Johnston and Dark, 1986; Egeth och Yantis, 1997). För att sammanfatta visar studier som visar effekter av uppmärksamhetskontroll uppifrån och ner att uppmärksamhet framgångsrikt kan tilldelas rumsliga platser, funktioner, objekt etc., efter närvaro av exogena eller endogena signaler (Eriksen och Hoffman, 1972; Posner, 1980; Posner et al., 1980), eller förväntningar antingen fastställda av förkunskaper eller av stimulansens händelser (Shaw, 1978; Moore och Egeth, 1998; Geng och Behrmann, 2002, 2005; Shomstein och Yantis, 2004A; Drummond och Shomstein, 2010). Bevis som stöder bottom-up attentional allocation har förlitat sig på olika attentional capture-paradigmer, där deltagarna är engagerade i en top-down-sökning och deras uppmärksamhet avleds till uppgiften-irrelevanta stimuli, vilket visar att uppmärksamhet fångas av feature singletons (unikt objekt; Yantis och Jonides, 1990; Theeuwes, 1991; Folk et al., 2002) och abrupta onsets (Yantis och Jonides, 1984; Theeuwes, 1991; Koshino et al., 1992; Juola et al., 1995).
medan de flesta tidiga studier koncentrerade sig på att visa bevis för uppmärksamhetsval uppifrån och ner, skiftade de senaste studierna fokus för att undersöka hur de två uppmärksamhetsvalssystemen interagerar. Denna undersökningslinje drivs av observationer som för att effektivt välja uppgiftsrelevant information (t.ex. ingredienser till salladen) måste man aktivt hämma uppgiftsrelaterad information som annars skulle avleda uppmärksamheten från uppgiften. Baksidan av denna logik är att ju mindre man fokuserar på uppgiftsrelaterad information, desto mer fångst kommer att uppstå. Det har visats experimentellt att observatörens uppmärksamhetstillstånd förutsäger vilken typ av information, och i vilken utsträckning, i slutändan kommer att fånga uppmärksamhet (Folk et al., 1992, 2002; Bacon och Egeth, 1994; Gibson och Kelsey, 1998). Till exempel Folk et al. (2002) visade att när man söker efter en röd bokstav kommer en observatör lättare att fångas av en irrelevant stimulans i periferin om den stimulansen är röd eller matchar målmallen på något sätt. Eftersom observatörens top-down-kontrollinställningar är inställda för att söka efter en röd funktion, kommer varje stimulans som är röd sannolikt att fånga uppmärksamhet och potentiellt störa top-down-kontrollen. Således, med en fångstuppgift, kan uppmärksamhetssökningsstrategier särskiljas från varandra genom att variera likhetsnivåerna mellan stimulusegenskaperna hos målet och distraktorerna. Ju mer liknande målet är för distraktorn, desto svårare är det för observatören att undvika fångst.
Parietallobens roll i valet uppifrån och ner och upp: Neuroimaging
olika neuroimaging tekniker gav starka bevis för involvering av parietal cortex i uppifrån och ner och uppåt orientering, med bevisen granskade i stor utsträckning någon annanstans (Corbetta och Shulman, 2002, 2011; Behrmann et al., 2004). Det har visats att områden som oftast aktiveras efter top-down-signaler för att delta i särskilda platser, funktioner eller föremål finns längs dorsala delar av parietal cortex. Sådana områden inkluderar underlägsen parietal lobule (IPL), dorsomediala regioner som kallas överlägsen parietal lobule (SPL), liksom mer mediala regioner längs precuneus gyrus (Yantis et al., 2002; Giesbrecht et al., 2003; Liu et al., 2003; Yantis och Serences, 2003; Figur 1). Flera top-down-uppgifter Har visats för att framgångsrikt engagera dorsala regioner i parietal cortex, nämligen de som involverar rumsliga (Kastner et al., 1999; Corbetta et al., 2000; Hopfinger et al., 2000; Shomstein och Behrmann, 2006; Chiu och Yantis, 2009; Greenberg et al., 2010) såväl som icke-rumsliga skift av uppmärksamhet (Giesbrecht et al., 2003; Yantis och Serences, 2003; Shomstein och Yantis, 2004b, 2006; Tamber-Rosenau et al., 2011).
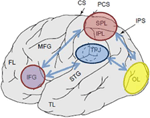
Figur 1. Schematisk bild av relevanta anatomiska landmärken projicerade på den mänskliga hjärnans laterala yta. Överlägsen parietal lobule (SPL) och inferior parietal lobule (IPL) är regioner inom den dorsala delen av parietal cortex som sänker uppifrån och ner uppmärksamhetsorientering. Temporo-parietal junction (TPJ) är en region inom den ventrala parietala cortex som sänker uppmärksamhetsorienteringen från botten upp. Både SPL och TPJ tros framkalla kontrollsignaler som är ansvariga för efterföljande uppmärksamhetsmoduleringar observerade över sensoriska regioner, i detta fall modulerande (märkt med mörkblå pilar) visuellt framkallad aktivitet i occipitalloben (OL). Dessutom tros områden längs inferior frontal gyrus (IFG) och inferior frontal junction (IFJ) fungera som konvergensområden för stimulansdriven och uppifrån och ner uppmärksamhetskontroll (markerad med ljusblå Dubbelriktade pilar).
i en typisk uppgift som syftar till att engagera uppifrån och ner uppmärksamhetsallokering visas individer två rapid serial visual presentation (RSVP) strömmar placerade perifert och instrueras initialt att övervaka en ström för en cue (t.ex. en siffra bland bokstavsströmmen). Identiteten på cue indikerar om ämnet måste behålla uppmärksamhet på den aktuella strömmen eller flytta uppmärksamheten till den andra strömmen (Yantis et al., 2002; Yantis och Serences, 2003). Två stora fynd observeras i sådana paradigmer. Den första har att göra med ökad aktivering inom de sensoriska regionerna som representerar den aktuella platsen (t.ex. ökad aktivitet inom de vänstra primära visuella regionerna när den högra RSVP-strömmen deltar). Detta resultat ger fasta bevis för att deltagarna deltar i en specifik plats och att uppmärksamhet modulerar styrkan i det sensoriska svaret (se Figur 1; Moran och Desimone, 1985; O ’ Craven et al., 1997). Det andra fyndet har att göra med observationen att dorsala regioner i parietalloben aktiveras selektivt genom skift av uppifrån och ner uppmärksamhet. Det observeras att SPL / IPL-tidsförloppet av aktivitet är övergående i naturen vilket tyder på att detta område av parietal cortex är källan till en kort uppmärksamhetskontrollsignal för att flytta uppmärksamma tillstånd på ett uppifrån och ner sätt (Yantis et al., 2002).
flera fMRI-studier har dokumenterat att bottom-up attentional capture, medierad av stimulus salience och/eller relevans, subserveras av temporo-parietal junction (TPJ; Figur 1). Till exempel, när ämnen deltar i och övervakar en förändring i antingen en visuell eller auditiv stimulans, presenteras samtidigt, förbättras aktiveringen av TPJ-regionerna i parietalloben. Förutom den uppenbara känsligheten för relevanta stimuli aktiveras TPJ också som svar på potentiellt nya (oväntade eller sällsynta) händelser när en organism är engagerad i ett neutralt beteendekontext eller när den är engagerad i en uppgift (Marois et al., 2000; Downar et al., 2002; Serences et al., 2005; Corbetta et al., 2008; Asplund et al., 2010; Diquattro och Geng, 2011; Geng och Mangun, 2011). Denna aktivering sker oberoende av modaliteten (auditiv, taktil och visuell) där ingången levereras, vilket återspeglar TPJ: s multisensoriska natur (men se Downar et al., 2001).
i en typisk uppgift som undersöker den neurala mekanismen för bottom-up attentional capture presenteras deltagarna med en RSVP-ström av objekt i mitten av displayen och uppmanas att identifiera ett fördefinierat mål (t.ex. identifiera röd bokstav som presenteras i en RSVP-ström av vita icke-mål). En del av försöken innehåller en uppgift-irrelevant framträdande distraktor som presenteras vid olika tidsintervall före målets början, medan andra försök endast innehåller den framträdande distraktorn (dvs. utan målet). ”Target-distractor” – försök används för att analysera omfattningen av fångst, vilket visar att den uppgift-irrelevanta distraktorn faktiskt är framträdande och därigenom ger en minskning av målnoggrannheten. ”Distractor-in-isolation” – försöken används för ytterligare analyser eftersom sådana försök möjliggör undersökning av aktivitet som framkallas till den framträdande distraktorn utan kontaminering från målrelaterade processer. Flera viktiga resultat framgår av sådana paradigmer. Först, när distraktorer separeras rumsligt från målplatsen, åtföljs fångstdistraktorer av ökad kortikal aktivitet i motsvarande regioner i sensorisk cortex (t.ex. retinotopiskt organiserad visuell cortex; se Figur 1). Sådana resultat ger starka bevis för att rumslig uppmärksamhet under fångst faktiskt fångas till den rumsliga platsen som upptas av distraktorn (Serences et al., 2005). För det andra aktiveras ventrala regioner i parietal cortex, huvudsakligen inom TPJ selektivt genom bottom-up, ofrivilliga, skift av uppmärksamhet. Precis som aktivitet inom SPL för orienteringen uppifrån och ner, är tidsförloppet för aktivitet observerad över TPJ övergående i naturen vilket tyder på att denna region är källan till en kort uppmärksamhetskontrollsignal för att flytta uppmärksamhet på ett bottom-up-sätt.
det bör noteras att även om denna översyn är inriktad på att ta itu med kognitiva funktioner i den bakre parietala cortexen, rekryteras andra regioner, särskilt de inom frontal cortex, också för uppmärksamhetsallokering uppifrån och ner. Sådana regioner inkluderar ventral frontal cortex (VFC), frontal eye fields (FEF), inferior frontal junction (IFJ) och inferior frontal gyrus (IFG; Corbetta och Shulman, 2002, 2011; Serences et al., 2005; Asplund et al., 2010; Diquattro och Geng, 2011).
Parietallobens roll i valet uppifrån och ner och upp: Neuropsykologi
historiskt förlitade sig forskare kritiskt på neuropsykologiska studier av patienter med hemispatial försummelse (en störning av rumslig fördelning av uppmärksamhet åt vänster hemi-utrymme) för att få insikt i kognitiva funktioner associerade med parietalloben. I den klassiska neuropsykologiska litteraturen ansågs parietal cortex som helhet allmänt vara den primära lesionsstället för hemispatial försummelse. Denna uppfattning, utarbetad i detalj av tidiga forskare (Critchley, 1953; McFire och Zangwill, 1960; Piercy, 1964) erkände tydligt sambandet mellan parietalskadan och den efterföljande försummelsen. Detta perspektiv hölls till stor del genom 1980-talet när Posner och kollegor (1984) använde det hemliga visuospatiala cueing-paradigmet för att visa att skador på parietalloben ger ett underskott i ”Koppla ur” – operationen (dra tillbaka uppmärksamhet från en plats och flytta den till en annan) när målet är kontralateralt mot lesionen. Trots detta stora framsteg när det gäller att förstå den neurala grunden för uppmärksamhet och specifikt parietal cortex ”urkoppling” Roll, antar deras resultat en enda kortikal plats (parietal cortex) och en enda funktionell förmåga (”urkoppling”). I motsats till detta mer monolitiska tillvägagångssätt för hjärnan (parietal cortex) och beteende (attentional disengagement), föreslår nyligen beteendemässigt och neuroimagingarbete (granskat ovan och på andra håll) att både den kortikala regionen och det associerade uppmärksamhetsbeteendet kan delas in i kvalitativt olika profiler.
Med tanke på segregering av de kortikala nätverken i top-down och bottom-up-processer är en uppenbar förutsägelse att skador på överlägsna delar av parietal lobule (subsuming SPL) bör ge ett underskott i målinriktad uppmärksamhetsorientering, medan skador på de sämre delarna av parietal lobule (subsuming TPJ) skulle resultera i ett underskott i samband med stimulansdriven uppmärksamhetsfångst. I den utsträckning som dessa hjärnbeteendekorrespondenser har utforskats i den neuropsykologiska litteraturen, är denna förutsägelse uppenbarligen inte upprätthållen. Till exempel är kliniska symtom på hemispatial försummelse starkt förknippade med skador på de sämre delarna av parietalloben, som inkluderar TPJ, snarare än till överlägsna delar som SPL (Friedrich et al., 1998; Shomstein et al., 2010; Corbetta och Shulman, 2011). Detta är något i strid med neuroimaging-litteraturen, vilket tyder på att TPJ: s roll är att fånga uppmärksamhet, snarare än i frivillig orientering av uppmärksamhet, den domän där försummelsespatienter verkar ha svårast. För att komplicera saken ytterligare har det noterats att lesioner som uteslutande involverar SPL endast sällan ger kliniska bevis på försummelse (Vallar och Perani, 1986). En annan ny studie med patienter med lesioner centrerade främst över TPJ och STG men bevarade SPL, Corbetta et al. (2005) visade att rumslig försummelse, liksom dess återhämtning, var förknippad med återställande av aktivitet i både ventrala temporo-parietala och dorsala parietala regioner (se Corbetta och Shulman, 2011 för en översyn). Även om det är intressant och spännande i sina slutsatser, skiljer den här sista studien inte det relativa bidraget från dorsala och ventrala vägar till olika typer av uppmärksamhet, eftersom patienter endast testades på en variant av Posner covert spatial attention cuing-uppgiften, uppgift som tros engagera både uppifrån och ner och upp uppmärksamhetsorientering.
för att skilja mellan måldriven uppmärksamhetskontroll och framträdande uppmärksamhetsfångst och att undersöka deras kartläggning på SPL respektive TPJ, antog ny studie två beteendeparadigmer, var och en riktade sig mot en av dessa former av uppmärksamhet (Shomstein et al., 2010). För att undersöka integriteten hos uppifrån och ner uppmärksamhetsorientering hos patienterna användes en uppifrån och ner-uppgift som krävde att deltagarna skiftade rumslig uppmärksamhet mellan de rumsligt separerade RSVP-strömmarna (en uppgift som framgångsrikt har använts för att demonstrera SPL-aktivering i fMRI-studier (Yantis et al., 2002)). Liknande, för att undersöka patientens uppmärksamhetsorienteringsförmåga, en variant av Folk et al. (2002) contingent capture paradigm användes där deltagarna upptäckte mål som uppträdde vid fixering medan uppgiftsrelaterade färg-singletoner blinkade i periferin. I vilken utsträckning uppgiftsrelaterade distraktorer stör den centrala detekteringsuppgiften användes sedan som ett mått på uppmärksamhetsfångst från botten upp (Bacon och Egeth, 1994; Folk et al., 2002).
förutsägelserna var följande: patienter med lesioner till överlägsna delar av parietalloben (som påverkar SPL) bör försämras i uppifrån och ner uppmärksamhetsorienteringsuppgiften (med bevarad prestanda på infångningsuppgiften) medan patienter med lesioner till de sämre delarna av parietalloben (som påverkar TPJ) bör försämras på infångningsuppgiften (med Sparad prestanda på uppifrån och ner-uppgiften). En dubbel dissociation av denna form intygar inte bara de oberoende komponenterna i uppmärksamhet utan antyder också att sådana uppmärksamhetskomponenter förmedlas av oberoende neurala mekanismer. Åtta patienter med visuo-rumslig försummelse rekryterades för studien och slutförde två uppgifter, knackade på antingen stimulansdriven eller målinriktad uppmärksamhetsorientering. Baserat på deras beteendeprofil sorterades patienterna i grupper och deras överlappning av lesionen undersöktes (figur 2a). Patienter som uppvisade svårigheter med målinriktad uppmärksamhetsorientering, som kvantifieras av top-down attentional index (Figur 2B), presenterade med lesion överlappning centrerad över överlägsna delar av parietal lobule (subsuming SPL) med Sparad inferior parietal lobule (TPJ). Patienter med lesion överlappar centrerad över de underlägsna delarna av parietal lobule (subsuming TPJ) men skonade SPL som utfördes normalt på den målinriktade orienteringsuppgiften, medan de förblev immuna mot uppmärksamhetsfångst (figur 2C). Resultaten från denna studie tyder tydligt på att SPL och TPJ är anatomiska regioner som nödvändigtvis rekryteras i syfte att orientera uppifrån och ner och uppåt och att skador på SPL och TPJ leder till störningar av orientering uppifrån och ner respektive.
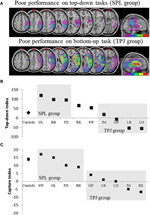
Figur 2. Resultat av neuropsykologisk studie som syftar till att undersöka det relativa bidraget från SPL och TPJ till uppifrån och ner och bottom-up orientering. (A) Lesion överlappar (lila minimal överlappning, röd maximal överlappning) för patienter grupperade efter beteendeunderskott i uppifrån och ner uppmärksamhetsorientering, märkt SPL-gruppen (toppanelen); och patienter grupperade efter beteendeunderskott i bottom-up-orientering, märkt TPJ-gruppen (nedre panelen). (B) Beteendeprestanda på top-down-uppgiften sammanfattad med ett ”Top-down Index” som kvantifierar skillnader mellan rumsliga top-down-Skift gjorda från vänster till höger och vice versa. Kontroller och den TPJ-skadade gruppen visar liknande effektivitet vid utförande av rumsliga Skift, medan patienter med SPL-lesioner visar minskad effektivitet. Gruppkontroll och individuella patientdata (märkta med patientinitialer) ritas på abscissen. (C)” Capture index ” är ett mått på bottom-up uppmärksamhet och kvantifierar i vilken utsträckning uppgift irrelevanta distraktorer fånga uppmärksamhet från uppgiften. Kontroller och spl-lesionerade gruppen visar liknande infångningsvärden, så att båda grupperna fångas av de uppgiftsrelaterade distraktorerna. TPJ lesioned group visar mycket reducerat fångstindex (misslyckande att fångas). Observera att patienter placerades i SPL-eller TPJ-gruppen baserat på beteende, snarare än baserat på lesionen, så notera konsistensen med vilken patienter hamnar i motsvarande grupp.
interaktion mellan top-Down och Bottom-Up Selection
även om det tydligen finns en stark koppling mellan målriktad orientering och SPL och stimulansdriven orientering och TPJ, data från Shomstein et al. (2010) patientstudie tyder på att dessa två system inte är helt oberoende. Denna slutsats stöds av upptäckten att patienter med SPL-skada uppvisade ett prestandamönster märkt som ”hyper capture.”Till skillnad från kontroller, för vilka endast målfärgad distraktor fångade uppmärksamhet (vilket ledde till lägre målnoggrannhet), visade sig irrelevanta färgade distraktorer också vara distraherande för patienter med SPL-lesion. För kontroller fångades uppmärksamhet endast av distraktorer när de föregick målet, för patienter med SPL-lesioner fångades uppmärksamhet till och med av distraktorer som presenterades samtidigt med målet. Detta prestandamönster kan förklaras med följande ramverk: SPL ansvarar för uppifrån och ner vägledning av uppmärksamhet som inkluderar att bestämma aspekterna av stimuli som är uppgift relevant (t.ex. Sök efter rött mål; Corbetta och Shulman, 2002; Serences et al., 2005). Denna uppmärksamhetsuppsättning begränsar sedan TPJ, så att fångstmekanismen som förmedlas av TPJ endast utlöses av uppgiften relevant information (t.ex. röda distraktorer som fångar uppmärksamhet och grå distraktorer som inte fångar uppmärksamhet när de söker efter ett rött mål). Frånvaron av SPL förhindrar upprättandet av en uppgift relevant attentional uppsättning och därmed någon stimulans, uppgift relevant eller inte, anses viktigt därför fånga uppmärksamhet (t.ex. uppgift irrelevant distraktor fånga uppmärksamhet för SPL-gruppen) urskillningslöst.
det har föreslagits att SPL och TPJ kan interagera på minst ett av två möjliga sätt. Den första möjligheten är att TPJ fungerar som ett varningssystem som upptäcker beteenderelevanta stimuli men saknar den höga rumsliga upplösningen, så när en beteenderelevant stimulans detekteras levereras dess exakta plats av SPL som lagrar rumsliga kartor (Kastner et al., 1999; Wojciulik och Kanwisher, 1999; Bisley och Goldberg, 2003; Silver et al., 2005). En relaterad hypotetisk möjlighet är att fångstmekanismen (som inkluderar TPJ) fungerar som en brytare av pågående kognitiv aktivitet när en beteendemässigt relevant stimulans presenteras (Corbetta och Shulman, 2002, 2011). ”Hyper-capture” aktivitetsmönstret observerat hos patienter med bevarad TPJ men lesionerad SPL ger ytterligare bevis för hypotesen att ser TPJ som utfärdande av en styrsignal som avslutar uppgiften till hands och därmed fungerar som en brytare (Corbetta och Shulman, 2002; Serences et al., 2005). Andra nya neuroimaging-studier som använder olika paradigmer har gett ytterligare bevis för ett interaktivt förhållande mellan uppifrån och ner och upp och ner uppmärksamhetsorientering, och därefter för förhållandet mellan SPL och TPJ (Serences et al., 2005; Asplund et al., 2010; Diquattro och Geng, 2011).
medan bevisen för en interaktion mellan de två uppmärksamhetssystemen och de två uppmärksamhetssubstraten (SPL och TPJ) är stark, är det fortfarande oklart om denna interaktion är direkt mellan SPL och TPJ eller om den uppnås genom andra mellanliggande regioner. Som nämnts tidigare, uppifrån och ner och upp uppmärksamhetsorienterande nätverk engagerar olika regioner inom frontal cortex, så det är rimligt att anta att konvergensen mellan de två systemen kan åstadkommas via frontalloben. Två nyligen genomförda studier som undersökte interaktionen mellan uppmärksamhetsval uppifrån och ner gav bevis för IFJ och IFG som möjliga konvergensplatser mellan stimulansdrivet och målriktat urval (Asplund et al., 2010; Diquattro och Geng, 2011). IFJ och IFG verkar vara idealiska kandidater för sådan interaktion med tanke på deras allmänna engagemang i uppmärksamhet och kognitiv kontroll samt dess engagemang i både rumsligt och icke-rumsligt urval (Koechlin et al., 2003; mässing et al., 2005).
Parietallobens roll i valet uppifrån och ner och upp: Fysiologi
medan tyngdpunkten i denna översyn huvudsakligen har lagts på mänskliga studier, har en stor mängd kunskap om involvering av parietal cortex i uppmärksamhetsorientering hämtats från monkey physiology investigations (se ny recension av Bisley och Goldberg, 2010). Men när det gäller att undersöka de relativa bidragen från olika regioner inom parietal cortex till uppifrån och ner och upp uppmärksamhetsorientering, faller apfysiologilitteraturen kort. Den främsta orsaken till detta är att inom apa cortex verkar det inte finnas bevis för samma segregering av top-down och bottom-up kontroll. Istället trodde lateralt intraparietalt område (LIP) ursprungligen att vara involverat i saccade-planering (Gnadt och Andersen, 1988) är involverat i visuell uppmärksamhet och fungerar som en prioriteringskarta där yttre stimuli representeras enligt deras beteendeprioritet härledda antingen uppifrån och ner eller nedifrån och upp (Colby och Goldberg, 1999; Bisley och Goldberg, 2003, 2010; Balan och Gottlieb, 2006; Ipata et al., 2006; Buschman och Miller, 2007; Gottlieb och Balan, 2010).
slutsats
Även om mycket mindre är känt om mänsklig parietal cortex än den för homolog apa cortex, har nya studier, som använder neuroimaging och neuropsykologiska metoder, börjat belysa alltmer finkorniga funktionella och strukturella skillnader. Denna översyn fokuserade på senaste neuroimaging och neuropsykologiska studier som belyser de kognitiva rollerna i dorsala och ventrala regioner i parietal cortex i uppifrån och ner och upp och ner uppmärksamhetsorientering, och på interaktionen mellan de två uppmärksamhetsallokeringsmekanismerna.
intressekonflikt uttalande
författaren förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
bekräftelser
detta arbete stöddes av National Institutes of Health grant EY021644.
Asplund, Cl, Todd, JJ, Snyder, AP och Marois, R. (2010). En central roll för den laterala prefrontala cortexen i målriktad och stimulansdriven uppmärksamhet. Nat. Neurovetenskap. 13, 507–512.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Bacon, WF och Egeth, he (1994). Övergripande stimulansdriven uppmärksamhetsfångst. Uppfattat. Psykofyser. 55, 485–496.
Pubmed Abstrakt / Pubmed fulltext
Balan, P. F., och Gottlieb, J. (2006). Integration av exogen input i en dynamisk salience karta avslöjas genom störande uppmärksamhet. J. Neurosci. 26, 9239–9249.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Behrmann, M., Geng, JJ och Shomstein, S. (2004). Parietal cortex och uppmärksamhet. Curr. Opin. Neurobiol. 14, 212–217.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Bisley, jw och Goldberg, Me (2003). Neuronal aktivitet i det laterala intraparietala området och rumslig uppmärksamhet. Vetenskap 299, 81-86.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Bisley, jw och Goldberg, Me (2010). Uppmärksamhet, avsikt och prioritet i parietalloben. Annu. Rev Neurosci. 33, 1–21.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
mässing, M., Derrfuss, J., Forstmann, B. Och von Cramon, D. Y. (2005). Rollen för det underlägsna frontala korsningsområdet i kognitiv kontroll. Trender Cogn. Sci. 9, 314–316.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Buschman, tj och Miller, ek (2007). Top-down kontra bottom-up kontroll av uppmärksamhet i prefrontala och bakre parietala cortices. Vetenskap 315, 1860-1862.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Chiu, YC och Yantis, S. (2009). En domänoberoende källa till kognitiv kontroll för uppgiftsuppsättningar: skiftande rumslig uppmärksamhet och byte av kategoriseringsregler. J. Neurosci. 29, 3930–3938.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Colby, Cl och Goldberg, Me (1999). Utrymme och uppmärksamhet i parietal cortex. Annu. Rev Neurosci. 22, 319–349.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Corbetta, M., Kincade, JM, Ollinger, JM, McAvoy, MP och Shulman, gl (2000). Frivillig orientering är dissocierad från måldetektering i mänsklig bakre parietal cortex. Nat. Neurovetenskap. 3, 292–297.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Corbetta, M., Kincade, MJ, Lewis, C., Snyder, AZ och Sapir, A. (2005). Neural grund och återhämtning av rumsliga uppmärksamhetsunderskott i rumslig försummelse. Nat. Neurovetenskap. 8, 1603–1610.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Corbetta, M., Patel, G. och Shulman, G. L. (2008). Den mänskliga hjärnans omorienteringssystem: från miljö till sinnesteori. Neuron 58, 306-324.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Corbetta, M. och Shulman, gl (2002). Kontroll av målinriktad och stimulansdriven uppmärksamhet i hjärnan. Nat. Rev Neurosci. 3, 201–215.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Corbetta, M. och Shulman, gl (2011). Rumslig försummelse och uppmärksamhetsnätverk. Annu. Rev Neurosci. 34, 569–599.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Critchley, M. (1953). Parietalloberna. London, Storbritannien: Hafner Press.
Pubmed Abstrakt / Pubmed fulltext
Diquattro, N. E. och Geng, J. J. (2011). Kontextuell kunskap konfigurerar uppmärksamhetskontrollnätverk. J. Neurosci. 31, 18026–18035.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Downar, J., Crawley, ap, Mikulis, DJ och Davis, KD (2001). Effekten av uppgiftsrelevans på det kortikala svaret på förändringar i visuella och auditiva stimuli: en händelserelaterad fMRI-studie. Neuroimage 14, 1256-1267.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Downar, J., Crawley, ap, Mikulis, DJ och Davis, KD (2002). Ett kortikalt nätverk som är känsligt för stimulans salience i ett neutralt beteendemässigt sammanhang över flera sensoriska modaliteter. J. Neurofysiol. 87, 615–620.
Pubmed Abstrakt / Pubmed fulltext
Drummond, L. och Shomstein, S. (2010). Objektbaserad uppmärksamhet: skiftande eller osäkerhet? Atten. Uppfattat. Psykofyser. 72, 1743–1755.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Egeth, He och Yantis, S. (1997). Visuell uppmärksamhet: kontroll, representation och tidskurs. Annu. Pastor Psychol. 48, 269–297.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Eriksen, CW och Hoffman, je (1972). Temporala och rumsliga egenskaper hos selektiv kodning från visuella skärmar. Uppfattat. Psykofyser. 12, 201–204.
Folk, Cl, Leber, AB och Egeth, he (2002). Fick dig att blinka! Contingent attentional capture ger en rumslig blinkning. Uppfattat. Psykofyser. 64, 741–753.
Pubmed Abstrakt / Pubmed fulltext
Folk, Cl, Remington, RW och Johnston, JC (1992). Ofrivillig hemlig orientering är beroende av uppmärksamhetskontrollinställningar. J. Exp. Psychol. Brum. Uppfattat. Utföra. 18, 1030–1044.
Pubmed Abstrakt / Pubmed fulltext
Friedrich, fj, Egly, R., Rafal, rd och Beck, D. (1998). Rumsliga uppmärksamhetsunderskott hos människor: en jämförelse av överlägsna parietala och temporala parietala korsningsskador. Neuropsykologi 12, 193-207.
Pubmed Abstrakt / Pubmed fulltext
Geng, JJ och Behrmann, M. (2002). Sannolikhet cuing av målplats underlättar visuell sökning implicit hos normala deltagare och patienter med hemispatial försummelse. Psychol. sci. 13, 520–525.
Pubmed Abstrakt / Pubmed fulltext
Geng, JJ och Behrmann, M. (2005). Spatial Sannolikhet som en uppmärksam cue i visuell sökning. Uppfattat. Psykofyser. 67, 1252–1268.
Pubmed Abstrakt / Pubmed fulltext
Geng, JJ och Mangun, gr (2011). Rätt temporoparietal korsningsaktivering av en framträdande kontextuell cue underlättar måldiskriminering. Neuroimage 54, 594-601.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Gibson, Bs och Kelsey, EM (1998). Stimulus driven attentional capture är beroende av attentional set för displaywide visuella funktioner. J. Exp. Psychol. Brum. Uppfattat. Utföra. 24, 699–706.
Pubmed Abstract/Pubmed fulltext
Giesbrecht, B., Woldorff, M. G., Song, A. W. och Mangun, G. R. (2003). Neurala mekanismer för top-down kontroll under rumslig och funktion uppmärksamhet. Neuroimage 19, 496-512.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Gnadt, jw och Andersen, ra (1988). Minnesrelaterad motorplaneringsaktivitet i bakre parietal cortex av makak. Exp. 70, 216-220.
Pubmed Abstrakt / Pubmed fulltext
Gottlieb, J. och Balan, P. (2010). Uppmärksamhet som ett beslut i informationsutrymme. Trender Cogn. Sci. 14, 240–248.
Pubmed Abstract / Pubmed fulltext / CrossRef fulltext
Greenberg, as, Esterman, M., Wilson, D., Serences, jt och Yantis, S. (2010). Kontroll av rumslig och funktionsbaserad uppmärksamhet i frontoparietal cortex. J. Neurosci. 30, 14330–14339.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Hopfinger, JB, Buonocore, MH och Mangun, gr (2000). De neurala mekanismerna för uppifrån och ner uppmärksamhetskontroll. Nat. Neurovetenskap. 3, 284–291.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Ipata, A. E., Gee, A. L., Gottlieb, J., Bisley, J. W. och Goldberg, M. E. (2006). LÄPPSVAR på en popout-stimulans reduceras om den ignoreras Öppet. Nat. Neurovetenskap. 9, 1071–1076.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Johnston, WA och Dark, VJ (1986). Selektiv Uppmärksamhet. Annu. Pastor Psychol. 37, 43–75.
Juola, JF, Koshino, H. och Warner, C. B. (1995). Avvägningar mellan attentional effekter av rumsliga signaler och abrupta onsets. Uppfattat. Psykofyser. 57, 333–342.
Pubmed Abstract / Pubmed fulltext
Kastner, S., Pinsk, M. A., De Weerd, P., Desimone, R. och Ungerleider, L. G. (1999). Ökad aktivitet i mänsklig visuell cortex under riktad uppmärksamhet i frånvaro av visuell stimulering. Neuron 22, 751-761.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Koechlin, E., Ody, C. och Kouneiher, F. (2003). Arkitekturen för kognitiv kontroll i den mänskliga prefrontala cortexen. Vetenskap 302, 1181-1185.
PubMed Abstrakt / Pubmed fulltext / CrossRef fulltext
Koshino, H., Warner, CB och Juola, JF (1992). Relativ effektivitet av centrala, perifera och abrupta signaler i visuell uppmärksamhet. Q. J. Exp. Psychol. En 45, 609-631.
Pubmed Abstrakt / Pubmed fulltext
Liu, T., Slotnick, SD, Serences, jt och Yantis, S. (2003). Kortikala mekanismer för funktionsbaserad uppmärksamhetskontroll. Cereb. Cortex 13, 1334-1343.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Marois, R., Leung, HC och Gore, JC (2000). Ett stimulansdrivet tillvägagångssätt för objektidentitet och platsbehandling i den mänskliga hjärnan. Neuron 25, 717-728.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
McFire, J. och Zangwill, O. L. (1960). Visuo-konstruktiva funktionshinder i samband med lesioner i vänstra hjärnhalvan. Hjärna 82, 243-260.
Moore, C. M. och Egeth, H. (1998). Hur påverkar funktionsbaserad uppmärksamhet visuell bearbetning? J. Exp. Psychol. Brum. Uppfattat. Utföra. 24, 1296–1310.
Pubmed Abstrakt / Pubmed fulltext
Moran, J. och Desimone, R. (1985). Selektiv uppmärksamhet grindar visuell bearbetning i extrastriate cortex. Vetenskap 229, 782-784.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
O ’ Craven, K., Rosen, B. R., Kwong, kk, Treisman, A. och Savoy, rl (1997). Frivillig uppmärksamhet modulerar fMRI-aktivitet i mänsklig MT-MST. Neuron 18, 591-598.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Piercy, M. (1964). Effekterna av hjärnskador på intellektuell funktion: en översyn av aktuella forskningstrender. Br. J. Psykiatri 110, 310-352.
Pubmed Abstrakt / Pubmed fulltext
Posner, M. I. (1980). Orientering av uppmärksamhet. Q. J. Exp. Psychol. 32, 3–25.
Pubmed Abstract / Pubmed fulltext
Posner, M. I., Snyder, C. R. och Davidson, B. J. (1980). Uppmärksamhet och detektering av signaler. J. Exp. Psychol. 109, 160–174.
Pubmed Abstract / Pubmed fulltext
Posner, M. I., Walker, J. A., Friedrich, F. J. och Rafal, R. D. (1984). Effekter av parietal skada på hemlig orientering av uppmärksamhet. J. Neurosci. 4, 1863–1874.
Pubmed Abstract / Pubmed fulltext
Serences, jt, Shomstein, S., Leber, AB, Golay, X., Egeth, He och Yantis, S. (2005). Samordning av frivillig och stimulansdriven uppmärksamhetskontroll i mänsklig cortex. Psychol. Sci. 16, 114–122.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Shaw, M. L. (1978). En kapacitetstilldelningsmodell för reaktionstid. J. Exp. Psychol. Brum. Uppfattat. Utföra. 4, 586–598.
Shomstein, S. och Behrmann, M. (2006). Kortikala system som förmedlar visuell uppmärksamhet åt både objekt och rumsliga platser. Proc. Natl. Acad. Sci. USA 103, 11387-11392.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Shomstein, S., Lee, J. och Behrmann, M. (2010). Top-down och bottom-up attentional vägledning: undersöka rollen av dorsala och ventrala parietala cortices. Exp. Hjärnan Res. 206, 197-208.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Shomstein, S. och Yantis, S. (2004A). Konfigurerbar och kontextuell prioritering i objektbaserad uppmärksamhet. Psykon. Tjur. Rev. 11, 247-253.
Pubmed Abstrakt / Pubmed fulltext
Shomstein, S. och Yantis, S. (2004b). Kontroll av uppmärksamhet skiftar mellan syn och audition i mänsklig cortex. J. Neurosci. 24, 10702–10706.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Shomstein, S. och Yantis, S. (2006). Parietal cortex förmedlar frivillig kontroll av rumslig och icke-rumslig auditiv uppmärksamhet. J. Neurosci. 26, 435–439.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Silver, M. A., Ress, D., och Heeger, D. J. (2005). Topografiska kartor över visuell rumslig uppmärksamhet i mänsklig parietal cortex. J. Neurofysiol. 94, 1358–1371.
Pubmed Abstrakt / Pubmed fulltext | CrossRef fulltext
Tamber-Rosenau, Bj, Esterman, M., Chiu, Y.-C. och Yantis, S. (2011). Kortikala mekanismer för kognitiv kontroll för att skifta uppmärksamhet i syn och arbetsminne. J. Cogn. Neurovetenskap. 23, 2905–2919.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Theeuwes, J. (1991). Exogen och endogen kontroll av uppmärksamhet: effekten av visuella onsets och offsets. Uppfattat. Psykofyser. 49, 83–90.
Pubmed Abstrakt / Pubmed fulltext
Vallar, G. och Perani, D. (1986). Anatomin av ensidig försummelse efter stroke på höger halvklot. En klinisk / CT-scan korrelationsstudie hos människa. Neuropsykologi 24, 609-622.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Wojciulik, E. och Kanwisher, N. (1999). Genera parietal engagemang i visuell uppmärksamhet. Neuron 23, 747-764.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Yantis, S. och Jonides, J. (1984). Abrupta visuella störningar och selektiv uppmärksamhet: bevis från visuell sökning. J. Exp. Psychol. Brum. Uppfattat. Utföra. 10, 601–621.
Pubmed Abstrakt / Pubmed fulltext
Yantis, S. och Jonides, J. (1990). Abrupta visuella uppkopplingar och selektiv uppmärksamhet: frivillig kontra automatisk tilldelning. J. Exp. Psychol. Brum. Uppfattat. Utföra. 16, 121–134.
Pubmed Abstrakt / Pubmed fulltext
Yantis, S., Schwarzbach, J., Serences, J. T., Carlson, R. L., Steinmetz, M. A., Pekar, J. J. och Courtney, S. M. (2002). Övergående neural aktivitet i mänsklig parietal cortex under rumsliga uppmärksamhetsskift. Nat. Neurovetenskap. 5, 995–1002.
Pubmed Abstrakt / Pubmed fulltext / CrossRef fulltext
Yantis, S. och Serences, J. T. (2003). Kortikala mekanismer för rymdbaserad och objektbaserad uppmärksamhetskontroll. Curr. Opin. Neurobiol. 13, 187–193.
PubMed Abstrakt / Pubmed fulltext / CrossRef fulltext