Einführung
Eine erfolgreiche Interaktion mit unserer sensorischen Umgebung erfordert ein kompliziertes Gleichgewicht zweier Aufmerksamkeitsauswahlmechanismen — dem von Top-Down und Bottom-up. Gehen Sie zum Produktgang Ihres örtlichen Supermarktes mit dem Ziel, nur wenige benötigte Zutaten für den Mangosalat aufzunehmen, und setzen Sie ein zielgerichtetes oder von oben nach unten gerichtetes Aufmerksamkeitssystem ein, sodass Sie aktiv nach allen erforderlichen Zutaten suchen die Vielzahl der Produktauswahl. Sollten Sie jedoch einen Klingelton eines Mobiltelefons hören, wird dies höchstwahrscheinlich Ihre Aufmerksamkeit erregen und Ihre Suche unterbrechen. Eine solche Unterbrechung erfolgt in einer Bottom-up- oder reizgesteuerten Weise, wobei eine bloße Hervorhebung des Reizes, die Tatsache, dass sich der Ring von anderen Geräuschen in Ihrer Umgebung unterscheidet, ihn für eine Auswahl würdig hält. Das beschriebene Szenario unterstreicht die Bedeutung einer zielgerichteten und stimulusgetriebenen Selektion für das Verhalten und weist auf ein feines Gleichgewicht hin, das zwischen den beiden Aufmerksamkeitssystemen bestehen muss, um „Tunnelblick“ einerseits und völlige Unfähigkeit, sich auf die andere zu konzentrieren, zu verhindern.
Auswahl von Oben nach unten und von unten nach oben: Verhalten
Mehrere Jahrzehnte der Verhaltensforschung haben gezeigt, dass die Verteilung der Aufmerksamkeit sowohl durch die Absichten des Beobachters als auch durch die Bedeutung des physischen Reizes gesteuert werden kann. Ein Großteil der Verhaltensbeweise für die Top-Down- und Bottom-Up-Aufmerksamkeitszuweisung wurde an anderer Stelle ausführlich überprüft (Johnston und Dark, 1986; Egeth und Yantis, 1997). Zusammenfassend zeigen Studien, die Effekte der Top-Down-Aufmerksamkeitskontrolle zeigen, dass Aufmerksamkeit erfolgreich räumlichen Orten, Merkmalen, Objekten usw. zugeordnet werden kann., nach Vorhandensein exogener oder endogener Hinweise (Eriksen und Hoffman, 1972; Posner, 1980; Posner et al., 1980) oder Erwartungen, die entweder durch Vorwissen oder durch Kontingenzen des Stimulus gesetzt werden (Shaw, 1978; Moore und Egeth, 1998; Geng und Behrmann, 2002, 2005; Shomstein und Yantis, 2004a; Drummond und Shomstein, 2010). Beweise für die Bottom-up-Aufmerksamkeitszuweisung stützten sich auf verschiedene Paradigmen zur Aufmerksamkeitserfassung, bei denen die Teilnehmer an einer Top-Down-Suche teilnehmen und ihre Aufmerksamkeit auf die aufgabenrelevanten Reize gelenkt wird, was zeigt, dass die Aufmerksamkeit von Merkmalssingletons erfasst wird (unique item; Yantis und Jonides, 1990; Theeuwes, 1991; Folk et al., 2002) und abrupte Einbrüche (Yantis und Jonides, 1984; Theeuwes, 1991; Koshino et al., 1992; Juola et al., 1995).Während sich die meisten frühen Studien darauf konzentrierten, Beweise für die Top-Down- und Bottom-Up-Aufmerksamkeitsauswahl zu demonstrieren, verlagerten die jüngsten Studien ihren Fokus auf die Untersuchung, wie die beiden Aufmerksamkeitsauswahlsysteme interagieren. Diese Untersuchungslinie wird durch Beobachtungen befeuert, dass man, um aufgabenrelevante Informationen (z. B. Zutaten für den Salat) effektiv auszuwählen, die aufgabenrelevanten Informationen aktiv hemmen muss, die sonst die Aufmerksamkeit von der vorliegenden Aufgabe ablenken würden. Die Kehrseite dieser Logik ist, dass je weniger man sich auf aufgabenbezogene Informationen konzentriert, desto mehr Erfassung erfolgt. Es wurde experimentell gezeigt, dass der Aufmerksamkeitszustand des Beobachters vorhersagt, welche Art von Information und in welchem Ausmaß letztendlich Aufmerksamkeit erregen wird (Folk et al., 1992, 2002; Speck und Egeth, 1994; Gibson und Kelsey, 1998). Zum Beispiel Folk et al. (2002) zeigten, dass ein Beobachter bei der Suche nach einem roten Buchstaben leichter von einem irrelevanten Stimulus in der Peripherie erfasst wird, wenn dieser Stimulus rot ist oder in irgendeiner Weise mit der Zielvorlage übereinstimmt. Da die Top-Down-Steuereinstellungen des Beobachters so eingestellt sind, dass sie nach einem roten Merkmal suchen, wird jeder rote Stimulus wahrscheinlich Aufmerksamkeit erregen und möglicherweise die Top-Down-Steuerung stören. So können bei einer Erfassungsaufgabe Aufmerksamkeitssuchstrategien voneinander unterschieden werden, indem die Ähnlichkeitsgrade zwischen den Reizeigenschaften des Ziels und den Distraktoren variiert werden. Je ähnlicher das Ziel dem Distraktor ist, desto schwieriger ist es für den Beobachter, die Erfassung zu vermeiden.
Die Rolle des Parietallappens bei der Top-Down- und Bottom-Up-Selektion: Neuroimaging
Verschiedene Neuroimaging-Techniken lieferten starke Beweise für die Beteiligung des parietalen Kortex an der Top-Down- und Bottom-up-Orientierung, wobei die Beweise an anderer Stelle ausführlich überprüft wurden (Corbetta und Shulman, 2002, 2011; Behrmann et al., 2004). Es wurde gezeigt, dass sich Bereiche, die am häufigsten nach Top-Down-Hinweisen aktiviert werden, um bestimmte Orte, Merkmale oder Objekte zu berücksichtigen, entlang der dorsalen Teile des parietalen Kortex befinden. Solche Bereiche umfassen den unteren Parietallappen (IPL), dorsomediale Regionen, die als oberer Parietallappen (SPL) bezeichnet werden, sowie medialere Regionen entlang des Gyrus precuneus (Yantis et al., 2002; Giesbrecht et al., 2003; Liu et al., 2003; Yantis und Serences, 2003; Abbildung 1). Es wurden mehrere Top-Down-Aufgaben entwickelt, um dorsale Regionen des parietalen Kortex erfolgreich zu aktivieren, nämlich diejenigen, die räumlich involviert sind (Kastner et al., 1999; Corbetta et al., 2000; Hopfinger et al., 2000; Shomstein und Behrmann, 2006; Chiu und Yantis, 2009; Greenberg et al., 2010) sowie nicht-räumliche Aufmerksamkeitsverschiebungen (Giesbrecht et al., 2003; Yantis und Serences, 2003; Shomstein und Yantis, 2004b, 2006; Tamber-Rosenau et al., 2011).
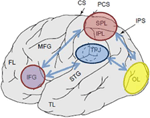
Abbildung 1. Schematische Darstellung relevanter anatomischer Landmarken projiziert auf die laterale Oberfläche des menschlichen Gehirns. Superior Parietal Lobule (SPL) und inferior Parietal Lobule (IPL) sind Regionen innerhalb des dorsalen Teils des Parietal Cortex, die eine Aufmerksamkeitsorientierung von oben nach unten ermöglichen. Der temporo-parietale Übergang (TPJ) ist eine Region innerhalb des ventralen parietalen Kortex, die eine Aufmerksamkeitsorientierung von unten nach oben ermöglicht. Es wird angenommen, dass beide, SPL und TPJ, Steuersignale hervorrufen, die für nachfolgende Aufmerksamkeitsmodulationen verantwortlich sind, die über sensorische Regionen beobachtet werden, in diesem Fall modulieren (markiert mit dunkelblauen Pfeilen) visuell evozierte Aktivität im Okzipitallappen (OL). Zusätzlich wird angenommen, dass Bereiche entlang des Gyrus frontalis inferior (IFG) und des Gyrus frontalis inferior (IFJ) als Konvergenzbereiche für die reizgesteuerte und von oben nach unten gerichtete Aufmerksamkeitskontrolle dienen (markiert durch hellblaue bidirektionale Pfeile).
In einer typischen Aufgabe, die darauf abzielt, die Aufmerksamkeitszuweisung von oben nach unten zu aktivieren, werden Einzelpersonen zwei RSVP-Streams (Rapid Serial Visual Presentation) gezeigt, die peripher positioniert sind, und sie werden zunächst angewiesen, einen Stream auf einen Hinweis zu überwachen (z. B. eine Ziffer unter dem Buchstabenstrom). Die Identität des Hinweises gibt an, ob das Subjekt die Aufmerksamkeit auf den aktuellen Strom richten oder die Aufmerksamkeit auf den anderen Strom lenken muss (Yantis et al., 2002; Yantis und Serences, 2003). Zwei wichtige Erkenntnisse werden in solchen Paradigmen beobachtet. Die erste hat mit einer erhöhten Aktivierung innerhalb der sensorischen Regionen zu tun, die den momentan besuchten Ort darstellen (z. B. erhöhte Aktivität innerhalb der linken primären visuellen Regionen, wenn der rechte RSVP-Stream besucht wird). Dieser Befund liefert einen festen Beweis dafür, dass die Teilnehmer an einem bestimmten Ort teilnehmen und dass die Aufmerksamkeit die Stärke der sensorischen Reaktion moduliert (siehe Abbildung 1; Moran und Desimone, 1985; O’Craven et al., 1997). Der zweite Befund hat mit der Beobachtung zu tun, dass dorsale Regionen des Parietallappens durch Verschiebungen der Top-Down-Aufmerksamkeit selektiv aktiviert werden. Es wird beobachtet, dass der SPL / IPL-Zeitverlauf der Aktivität vorübergehender Natur ist, was darauf hindeutet, dass dieser Bereich des parietalen Kortex die Quelle eines kurzen Aufmerksamkeitssteuersignals ist, um aufmerksame Zustände von oben nach unten zu verschieben (Yantis et al., 2002).
Mehrere fMRT-Studien haben dokumentiert, dass Bottom-up-Aufmerksamkeitserfassung, vermittelt durch Stimulus Salienz und / oder Relevanz, wird durch den temporo-parietalen Übergang (TPJ; Abbildung 1) subserviert. Wenn beispielsweise Probanden eine Änderung entweder eines visuellen oder eines auditiven Reizes gleichzeitig beobachten und überwachen, wird die Aktivierung der TPJ-Regionen des Parietallappens verstärkt. Zusätzlich zur offensichtlichen Empfindlichkeit gegenüber relevanten Reizen wird TPJ auch als Reaktion auf potenziell neuartige (unerwartete oder seltene) Ereignisse aktiviert, wenn ein Organismus in einem neutralen Verhaltenskontext oder bei einer Aufgabe tätig ist (Marois et al., 2000; Downar et al., 2002; Serences et al., 2005; Corbetta et al., 2008; Asplund et al., 2010; Diquattro und Geng, 2011; Geng und Mangun, 2011). Diese Aktivierung erfolgt unabhängig von der Modalität (auditiv, taktil und visuell), in der die Eingabe geliefert wird, was die multisensorische Natur von TPJ widerspiegelt (siehe jedoch Downar et al., 2001).In einer typischen Aufgabe, die den neuronalen Mechanismus der Bottom-up-Aufmerksamkeitserfassung untersucht, wird den Teilnehmern ein RSVP-Stream von Elementen in der Mitte des Displays präsentiert und sie werden gebeten, ein vordefiniertes Ziel zu identifizieren (z. B. Identifizieren Sie einen roten Buchstaben, der in einem RSVP-Stream von weißen Nichtzielen dargestellt wird). Ein Teil der Studien enthält einen aufgabenrelevanten hervorstechenden Distraktor, der in verschiedenen Zeitintervallen vor dem Einsetzen des Ziels präsentiert wird, während andere Studien nur den hervorstechenden Distraktor enthalten (d. H. ohne das Ziel). „Ziel-Distraktor“ -Versuche werden verwendet, um das Ausmaß der Erfassung zu bestimmen, was zeigt, dass der aufgabenrelevante Distraktor tatsächlich hervorstechend ist, was zu einer Abnahme der Zielgenauigkeit führt. Die „Distractor-in-Isolation“ -Versuche werden für weitere Analysen verwendet, da solche Versuche die Untersuchung der Aktivität ermöglichen, die dem hervorstechenden Distraktor ohne Kontamination durch die zielbezogenen Prozesse entlockt wird. Aus solchen Paradigmen ergeben sich mehrere wichtige Erkenntnisse. Erstens, wenn Distraktoren räumlich vom Zielort getrennt sind, werden Capture-Distraktoren von einer erhöhten kortikalen Aktivität in entsprechenden Regionen des sensorischen Kortex begleitet (z. B. retinotopisch organisierter visueller Kortex; siehe Abbildung 1). Solche Ergebnisse liefern einen starken Beweis dafür, dass während der Erfassung die räumliche Aufmerksamkeit tatsächlich auf den vom Distraktor eingenommenen räumlichen Ort gerichtet ist (Serences et al., 2005). Zweitens werden ventrale Regionen des parietalen Kortex, hauptsächlich innerhalb des TPJ, selektiv durch unwillkürliche Aufmerksamkeitsverschiebungen von unten nach oben aktiviert. Genau wie die Aktivität innerhalb des SPL für die Top-Down-Orientierung ist der über TPJ beobachtete zeitliche Verlauf der Aktivität vorübergehender Natur, was darauf hindeutet, dass diese Region die Quelle eines kurzen Aufmerksamkeitssteuersignals ist, um die Aufmerksamkeit von unten nach oben zu verlagern.Es sollte angemerkt werden, dass, während diese Überprüfung auf die Adressierung der kognitiven Funktionen des posterioren parietalen Kortex konzentriert ist, andere Regionen, insbesondere diejenigen innerhalb des frontalen Kortex, auch für die Top-Down- und Bottom-up-Aufmerksamkeitszuweisung rekrutiert werden. Solche Regionen umfassen den ventralen frontalen Kortex (VFC), die frontalen Augenfelder (FEF), den inferioren frontalen Übergang (IFJ) und den inferioren frontalen Gyrus (IFG; Corbetta und Shulman, 2002, 2011; Serences et al., 2005; Asplund et al., 2010; Diquattro und Geng, 2011).
Die Rolle des Parietallappens bei der Top-Down- und Bottom-Up-Selektion: Neuropsychologie
Die Forscher stützten sich kritisch auf neuropsychologische Studien von Patienten mit hemispatialer Vernachlässigung (eine Störung der räumlichen Verteilung der Aufmerksamkeit auf den linken Hemiraum), um Einblicke in kognitive Funktionen zu erhalten, die mit dem Parietallappen verbunden sind. In der klassischen neuropsychologischen Literatur wurde der parietale Kortex als Gesamtheit allgemein als primäre Läsionsstelle für hemispatiale Vernachlässigung angesehen. Diese Ansicht wurde von frühen Forschern ausführlich ausgearbeitet (Critchley, 1953; McFire und Zangwill, 1960; Piercy, 1964) erkannte deutlich den Zusammenhang zwischen der parietalen Läsion und der daraus resultierenden Vernachlässigung. Diese Perspektive wurde weitgehend durch die 1980er Jahre gehalten, als Posner und Kollegen (1984) das verdeckte Visuospatial Cueing Paradigma verwendeten, um zu zeigen, dass eine Schädigung des Parietallappens ein Defizit in der „Disengage“ -Operation (Zurückziehen der Aufmerksamkeit von einem Ort und Verlagerung auf einen anderen) erzeugt, wenn das Ziel kontralateral zur Läsion ist. Trotz dieses großen Fortschritts beim Verständnis der neuronalen Basis der Aufmerksamkeit und insbesondere der „Disengage“ -Rolle des parietalen Kortex gehen ihre Ergebnisse von einer einzigen kortikalen Stelle (parietaler Kortex) und einer einzigen Funktionsfähigkeit („Disengage“) aus. Im Gegensatz zu diesem monolithischeren Ansatz für das Gehirn (parietaler Kortex) und das Verhalten (Aufmerksamkeitsentkopplung) legen neuere Verhaltens- und Neuroimaging-Arbeiten (oben und anderswo überprüft) nahe, dass sowohl die kortikale Region als auch das damit verbundene Aufmerksamkeitsverhalten in qualitativ unterschiedliche Profile unterteilt werden können.Angesichts der Segregation der kortikalen Netzwerke in Top-Down- und Bottom-up-Prozesse ist eine offensichtliche Vorhersage, dass eine Schädigung der oberen Teile des Parietallappens (Subsuming SPL) zu einem Defizit bei der zielgerichteten Aufmerksamkeitsorientierung führen sollte, während eine Schädigung der unteren Teile des Parietallappens (Subsuming TPJ) zu einem Defizit führen würde, das mit einer stimulusgesteuerten Aufmerksamkeitserfassung verbunden ist. In dem Maße, in dem diese Gehirn-Verhaltens-Korrespondenzen in der neuropsychologischen Literatur untersucht wurden, wird diese Vorhersage offensichtlich nicht bestätigt. Zum Beispiel sind klinische Symptome einer hemispatialen Vernachlässigung stark mit einer Schädigung der unteren Teile des Parietallappens verbunden, die TPJ einschließt, und nicht mit überlegenen Teilen wie SPL (Friedrich et al., 1998; Schomstein et al., 2010; Corbetta und Shulman, 2011). Dies steht im Widerspruch zur Neuroimaging-Literatur, die darauf hindeutet, dass die Rolle von TPJ eher in der Erfassung der Aufmerksamkeit als in der freiwilligen Ausrichtung der Aufmerksamkeit liegt, dem Bereich, in dem Vernachlässigungspatienten die größten Schwierigkeiten zu haben scheinen. Um die Sache noch komplizierter zu machen, wurde festgestellt, dass Läsionen, die ausschließlich SPL betreffen, nur selten klinische Hinweise auf Vernachlässigung liefern (Vallar und Perani, 1986). Eine weitere kürzlich durchgeführte Studie mit Patienten mit Läsionen, die sich hauptsächlich auf TPJ und STG konzentrierten, aber SPL konservierten, Corbetta et al. (2005) zeigten, dass räumliche Vernachlässigung sowie ihre Wiederherstellung mit der Wiederherstellung der Aktivität sowohl in den ventralen temporo-parietalen als auch in den dorsalen parietalen Regionen verbunden waren (siehe Corbetta und Shulman, 2011 für eine Überprüfung). Obwohl diese letzte Studie in ihren Schlussfolgerungen interessant und aufregend ist, unterscheidet sie nicht den relativen Beitrag der dorsalen und ventralen Bahnen zu verschiedenen Arten von Aufmerksamkeit, da die Patienten nur an einer Variante der Posner-verdeckten räumlichen Aufmerksamkeits-Cuing-Aufgabe getestet wurden.
Um zwischen zielgerichteter Aufmerksamkeitskontrolle und auffälliger Aufmerksamkeitserfassung zu unterscheiden und ihre Zuordnung zum SPL bzw. TPJ zu untersuchen, wurden kürzlich zwei Verhaltensparadigmen verwendet, die jeweils auf eine dieser Formen der Aufmerksamkeit abzielen (Shomstein et al., 2010). Um die Integrität der Top-Down-Aufmerksamkeitsorientierung bei den Patienten zu untersuchen, wurde eine Top-Down-Aufgabe verwendet, bei der die Teilnehmer die räumliche Aufmerksamkeit zwischen den räumlich getrennten RSVP-Strömen verlagern mussten (eine Aufgabe, die erfolgreich zum Nachweis der SPL-Aktivierung in fMRT-Studien eingesetzt wurde (Yantis et al., 2002)). Ähnlich, um die Bottom-up-Aufmerksamkeitsorientierungsfähigkeiten der Patienten zu untersuchen, eine Variante von Folk et al. (2002) Es wurde ein kontingentes Erfassungsparadigma angewendet, bei dem die Teilnehmer Ziele erkannten, die bei der Fixierung auftraten, während aufgabenrelevante Farbsingletons in der Peripherie geflasht wurden. Das Ausmaß, in dem task-irrelevante Distraktoren die zentrale Erkennungsaufgabe stören, wurde dann als Maß für die Bottom-up-Aufmerksamkeitserfassung verwendet (Bacon und Egeth, 1994; Folk et al., 2002).
Die Vorhersagen waren wie folgt: patienten mit Läsionen an den oberen Teilen des Parietallappens (SPL beeinflussend) sollten bei der Top-Down-Aufmerksamkeitsorientierungsaufgabe beeinträchtigt sein (mit erhaltener Leistung bei der Erfassungsaufgabe), während Patienten mit Läsionen an den unteren Teilen des Parietallappens (TPJ beeinflussend) bei der Erfassungsaufgabe beeinträchtigt sein sollten (mit erhaltener Leistung bei der Top-Down-Aufgabe). Eine doppelte Dissoziation dieser Form bestätigt nicht nur die unabhängigen Komponenten der Aufmerksamkeit, sondern legt auch nahe, dass solche Aufmerksamkeitskomponenten durch unabhängige neuronale Mechanismen vermittelt werden. Acht Patienten mit visuell-räumlicher Vernachlässigung wurden für die Studie rekrutiert und absolvierten zwei Aufgaben, entweder stimulus-getriebene oder zielgerichtete Aufmerksamkeitsorientierung. Basierend auf ihrem Verhaltensprofil wurden die Patienten in Gruppen eingeteilt und ihre Läsionsüberlappung untersucht (Abbildung 2A). Patienten, die Schwierigkeiten mit der zielgerichteten Aufmerksamkeitsorientierung aufwiesen, wie durch den Top-Down-Aufmerksamkeitsindex quantifiziert (Abbildung 2B), zeigten eine Läsionsüberlappung, die über den oberen Teilen des Parietallappens (Subsuming SPL) mit dem unteren Parietallappen (TPJ) zentriert war. Patienten mit Läsionsüberlappung zentriert über die unteren Teile des Parietallappens (Subsuming TPJ), aber verschont SPL durchgeführt normalerweise auf die zielgerichtete Orientierungsaufgabe, während sie immun gegen Aufmerksamkeitserfassung bleiben (Abbildung 2C). Die Ergebnisse dieser Studie legen eindeutig nahe, dass SPL und TPJ anatomische Regionen sind, die notwendigerweise für die Zwecke der Top-Down- und Bottom-Up-Orientierung rekrutiert werden und dass Schäden an SPL und TPJ zu Störungen der Top-Down- bzw. Bottom-Up-Orientierung führen.
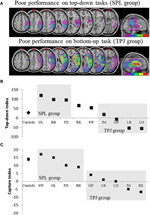
Abbildung 2. Ergebnisse der neuropsychologischen Studie zur Untersuchung des relativen Beitrags von SPL und TPJ zur Top-Down- und Bottom-Up-Orientierung. (A) Läsionsüberlappungen (lila minimale Überlappung; rote maximale Überlappung) für Patienten, die nach Verhaltensdefiziten in der Top-Down-Aufmerksamkeitsorientierung gruppiert sind, die als SPL-Gruppe (oberes Bedienfeld) bezeichnet wird; und Patienten, die nach Verhaltensdefiziten in der Bottom-up-Orientierung gruppiert sind, die als TPJ-Gruppe (unteres Bedienfeld) bezeichnet werden. (B) Verhaltensleistung bei der Top-Down-Aufgabe, zusammengefasst mit einem „Top-Down-Index“, der Unterschiede zwischen räumlichen Top-Down-Verschiebungen von links nach rechts und umgekehrt quantifiziert. Kontrollen und die TPJ-Läsionsgruppe zeigen ähnliche Effizienzen bei der Ausführung räumlicher Verschiebungen, während Patienten mit SPL-Läsionen eine verringerte Effizienz aufweisen. Auf der Abszisse sind Gruppenkontroll- und individuelle Patientendaten (mit Patienteninitialen gekennzeichnet) aufgetragen. (C) „Erfassungsindex“ ist ein Maß für die Bottom-up-Aufmerksamkeit und quantifiziert das Ausmaß, in dem aufgabenrelevante Distraktoren die Aufmerksamkeit von der Aufgabe ablenken. Kontrollen und die SPL-läsionierte Gruppe zeigen ähnliche Erfassungswerte, so dass beide Gruppen von den aufgabenrelevanten Distraktoren erfasst werden. TPJ-läsionierte Gruppen zeigen einen stark reduzierten Erfassungsindex (nicht erfasst werden). Beachten Sie, dass Patienten basierend auf dem Verhalten und nicht basierend auf der Läsion in die SPL- oder TPJ-Gruppe eingeteilt wurden, und beachten Sie daher die Konsistenz, mit der Patienten in die entsprechende Gruppe gelangen.
Interaktion zwischen Top-Down- und Bottom-Up-Selektion
Obwohl es anscheinend eine starke Assoziation zwischen zielgerichteter Orientierung und SPL und stimulusgetriebener Orientierung und TPJ gibt, zeigen Daten von Shomstein et al. (2010) Patientenstudie legen nahe, dass diese beiden Systeme nicht völlig unabhängig sind. Diese Schlussfolgerung wird durch die Feststellung gestützt, dass Patienten mit SPL-Schäden ein als „Hyper Capture“ bezeichnetes Leistungsmuster aufwiesen.“ Im Gegensatz zu Kontrollen, bei denen nur zielfarbige Distraktoren die Aufmerksamkeit erregten (was zu einer geringeren Zielgenauigkeit führte), erwiesen sich irrelevante farbige Distraktoren auch als störend für Patienten mit SPL-Läsion. Während bei Kontrollen die Aufmerksamkeit nur dann von Distraktoren erfasst wurde, wenn sie dem Einsetzen des Ziels vorausgingen, wurde bei Patienten mit SPL-Läsionen die Aufmerksamkeit sogar von Distraktoren erfasst, die gleichzeitig mit dem Ziel präsentiert wurden. Dieses Leistungsmuster kann durch das folgende Framework erklärt werden: SPL ist verantwortlich für die Top-Down-Führung der Aufmerksamkeit, die die Bestimmung der Aspekte der Stimuli umfasst, die aufgabenrelevant sind (z. B. Suche nach einem Ziel; Corbetta und Shulman, 2002; Serences et al., 2005). Dieser Aufmerksamkeitssatz schränkt dann TPJ ein, so dass der durch TPJ vermittelte Aufmerksamkeitserfassungsmechanismus nur durch die aufgabenrelevanten Informationen ausgelöst wird (z. B. rote Distraktoren, die Aufmerksamkeit erfassen, und graue Distraktoren, die bei der Suche nach einem roten Ziel keine Aufmerksamkeit erfassen). Das Fehlen von SPL verhindert die Etablierung eines aufgabenrelevanten Aufmerksamkeitssatzes, und daher wird jeder Stimulus, ob aufgabenrelevant oder nicht, als wichtig erachtet, um die Aufmerksamkeit wahllos zu erfassen (z. B. aufgabenrelevanter Distraktor, der die Aufmerksamkeit für die SPL-Gruppe erfasst).
Es wurde vorgeschlagen, dass SPL und TPJ auf mindestens eine von zwei möglichen Arten interagieren könnten. Die erste Möglichkeit besteht darin, dass TPJ als Warnsystem dient, das verhaltensrelevante Reize erkennt, jedoch nicht über die hohe räumliche Auflösung verfügt, sodass bei Erkennung eines verhaltensrelevanten Reizes dessen genaue Position von der SPL geliefert wird, die räumliche Karten speichert (Kastner et al., 1999; Wojciulik und Kanwisher, 1999; Bisley und Goldberg, 2003; Silber et al., 2005). Eine verwandte hypothetische Möglichkeit besteht darin, dass der Erfassungsmechanismus (einschließlich TPJ) als Leistungsschalter für die laufende kognitive Aktivität fungiert, wenn ein verhaltensrelevanter Stimulus präsentiert wird (Corbetta und Shulman, 2002, 2011). Das „Hyper-Capture“ -Aktivitätsmuster, das bei Patienten mit erhaltenem TPJ, aber läsioniertem SPL beobachtet wurde, liefert weitere Beweise für die Hypothese, dass TPJ ein Steuersignal ausgibt, das die vorliegende Aufgabe beendet und somit als Leistungsschalter dient (Corbetta und Shulman, 2002; Serences et al., 2005). Andere neuere Neuroimaging-Studien, die verschiedene Paradigmen verwenden, haben weitere Beweise für eine interaktive Beziehung zwischen der Top-Down- und der Bottom-Up-Aufmerksamkeitsorientierung und anschließend für die Beziehung zwischen SPL und TPJ geliefert (Serences et al., 2005; Asplund et al., 2010; Diquattro und Geng, 2011).Während die Beweise für eine Wechselwirkung zwischen den beiden Aufmerksamkeitssystemen und den beiden Aufmerksamkeitssubstraten (SPL und TPJ) stark sind, bleibt unklar, ob diese Wechselwirkung direkt zwischen SPL und TPJ stattfindet oder ob sie durch andere Zwischenregionen erreicht wird. Wie bereits erwähnt, greifen Top-Down- und Bottom-Up-Aufmerksamkeitsorientierungsnetzwerke verschiedene Regionen innerhalb des frontalen Kortex an, daher ist es vernünftig anzunehmen, dass die Konvergenz zwischen den beiden Systemen über den Frontallappen erreicht werden könnte. Zwei kürzlich durchgeführte Studien, die die Interaktion zwischen Top-Down- und Bottom-Up-Aufmerksamkeitsauswahl untersuchten, lieferten Hinweise auf IFJ und IFG als mögliche Konvergenzstellen zwischen stimulusgetriebener und zielgerichteter Selektion (Asplund et al., 2010; Diquattro und Geng, 2011). Das IFJ und IFG scheinen ideale Kandidaten für eine solche Interaktion zu sein, da sie allgemein an Aufmerksamkeit und kognitiver Kontrolle sowie an räumlicher und nicht räumlicher Selektion beteiligt sind (Koechlin et al., 2003; Brass et al., 2005).
Die Rolle des Parietallappens bei der Top-Down- und Bottom-Up-Selektion: Physiologie
Während der Schwerpunkt dieser Überprüfung überwiegend auf Humanstudien gelegt wurde, wurde aus Untersuchungen zur Affenphysiologie ein großer Wissensschatz über die Beteiligung des parietalen Kortex an der Aufmerksamkeitsorientierung gewonnen (siehe jüngste Überprüfung von Bisley und Goldberg, 2010). Wenn es jedoch darum geht, die relativen Beiträge verschiedener Regionen innerhalb des parietalen Kortex zur Aufmerksamkeitsorientierung von oben nach unten und von unten nach oben zu untersuchen, greift die Literatur zur Affenphysiologie zu kurz. Der Hauptgrund dafür ist, dass es innerhalb des Affenkortex keine Beweise für die gleiche Trennung von Top-Down- und Bottom-Up-Kontrolle zu geben scheint. Stattdessen ist der laterale intraparietale Bereich (LIP), von dem ursprünglich angenommen wurde, dass er an der Sakkadenplanung beteiligt ist (Gnadt und Andersen, 1988), an der visuellen Aufmerksamkeit beteiligt und fungiert als Prioritätskarte, in der externe Reize entsprechend ihrer Verhaltenspriorität dargestellt werden, die entweder von oben nach unten oder von unten nach oben abgeleitet ist (Colby und Goldberg, 1999; Bisley und Goldberg, 2003, 2010; Balan und Gottlieb, 2006; Ipata et al., 2006; Buschman und Miller, 2007; Gottlieb und Balan, 2010).
Schlussfolgerung
Obwohl viel weniger über den menschlichen Parietalcortex bekannt ist als über den homologen Affencortex, haben neuere Studien, die bildgebende und neuropsychologische Methoden anwenden, begonnen, zunehmend feinkörnige funktionelle und strukturelle Unterschiede aufzuklären. Diese Übersicht konzentrierte sich auf neuere Neuroimaging- und neuropsychologische Studien, die die kognitiven Rollen der dorsalen und ventralen Regionen des parietalen Kortex bei der Top-Down- und Bottom-up-Aufmerksamkeitsorientierung sowie die Interaktion zwischen den beiden Aufmerksamkeitszuordnungsmechanismen aufklären.
Erklärung zum Interessenkonflikt
Der Autor erklärt, dass die Forschung in Abwesenheit von kommerziellen oder finanziellen Beziehungen durchgeführt wurde, die als potenzieller Interessenkonflikt ausgelegt werden könnten.
Danksagung
Diese Arbeit wurde vom National Institutes of Health Grant EY021644 unterstützt.
Asplund, C. L., Todd, J. J., Snyder, A. P. und Marois, R. (2010). Eine zentrale Rolle für den lateralen präfrontalen Kortex bei der zielgerichteten und reizgesteuerten Aufmerksamkeit. Nat. Neurowissenschaften. 13, 507–512.
Pubmed Abstract | Pubmed Full Text / CrossRef Full Text
Speck, W. F., und Egeth, H. E. (1994). Überschreiben der reizgesteuerten Aufmerksamkeitserfassung. Wahrnehmung. In: Psychophys. 55, 485–496.
Pubmed Zusammenfassung / Pubmed Volltext
Balan, P. F., und Gottlieb, J. (2006). Integration von exogenem Input in eine dynamische Salienzkarte, die durch störende Aufmerksamkeit aufgedeckt wird. In: J. Neurosci. 26, 9239–9249.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Behrmann, M., Geng, J. J. und Shomstein, S. (2004). Parietal Cortex und Aufmerksamkeit. Curr. Opin. Neurobiol. 14, 212–217.
Pubmed Zusammenfassung / Pubmed Volltext / CrossRef Volltext
Bisley, JW und Goldberg, ME (2003). Neuronale Aktivität im lateralen intraparietalen Bereich und räumliche Aufmerksamkeit. Wissenschaft 299, 81-86.
Pubmed Zusammenfassung / Pubmed Volltext / CrossRef Volltext
Bisley, JW und Goldberg, ME (2010). Aufmerksamkeit, Absicht und Priorität im Parietallappen. Jahr. Rev. Neurosci. 33, 1–21.
Pubmed Abstract | Pubmed Full Text / CrossRef Full Text
Brass, M., Derrfuss, J., Forstmann, B. und von Cramon, D. Y. (2005). Die Rolle des inferioren Frontalknotenbereichs bei der kognitiven Kontrolle. Trends Cogn. Sci. 9, 314–316.
Pubmed Zusammenfassung / Pubmed Volltext / CrossRef Volltext
Buschman, TJ und Miller, EK (2007). Top-down versus Bottom-up Kontrolle der Aufmerksamkeit in den präfrontalen und posterioren parietalen Kortizes. Wissenschaft 315, 1860-1862.
Pubmed Zusammenfassung / Pubmed Volltext / CrossRef Volltext
Chiu, YC und Yantis, S. (2009). Eine domänenunabhängige Quelle kognitiver Kontrolle für Aufgabensätze: Verschiebung der räumlichen Aufmerksamkeit und Wechsel der Kategorisierungsregeln. In: J. Neurosci. 29, 3930–3938.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
Colby, C. L. und Goldberg, M. E. (1999). Raum und Aufmerksamkeit im parietalen Cortex. Jahr. Rev. Neurosci. 22, 319–349.
Pubmed Abstract | Pubmed Full Text / CrossRef Full Text
Corbetta, M., Kincade, J. M., Ollinger, J. M., McAvoy, M. P. und Shulman, G. L. (2000). Freiwillige Orientierung ist von der Zieldetektion im menschlichen posterioren parietalen Kortex dissoziiert. Nat. Neurowissenschaften. 3, 292–297.
Pubmed Abstract | Pubmed Full Text / CrossRef Full Text
Corbetta, M., Kincade, M. J., Lewis, C., Snyder, A. Z. und Sapir, A. (2005). Neuronale Basis und Wiederherstellung räumlicher Aufmerksamkeitsdefizite bei räumlicher Vernachlässigung. Nat. Neurowissenschaften. 8, 1603–1610.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
Corbetta, M., Patel, G. und Shulman, GL (2008). Das Neuorientierungssystem des menschlichen Gehirns: von der Umwelt zur Theorie des Geistes. Neuron 58, 306-324.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
Corbetta, M. und Shulman, GL (2002). Kontrolle der zielgerichteten und reizgesteuerten Aufmerksamkeit im Gehirn. Nat. Rev. Neurosci. 3, 201–215.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
Corbetta, M. und Shulman, GL (2011). Spatial Neglect und Aufmerksamkeitsnetzwerke. Jahr. Rev. Neurosci. 34, 569–599.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
Critchley, M. (1953). Die Parietallappen. London, Großbritannien: Hafner Press.
Pubmed Zusammenfassung / Pubmed Volltext
Diquattro, N. E. und Geng, J. J. (2011). Kontextwissen konfiguriert Aufmerksamkeitssteuerungsnetzwerke. In: J. Neurosci. 31, 18026–18035.
Pubmed Zusammenfassung / Pubmed Volltext / CrossRef Volltext
Downar, J., Crawley, AP, Mikulis, DJ und Davis, KD (2001). Die Auswirkung der Aufgabenrelevanz auf die kortikale Reaktion auf Veränderungen visueller und auditiver Reize: eine ereignisbezogene fMRT-Studie. Neuroimage 14, 1256-1267.
Pubmed Zusammenfassung / Pubmed Volltext / CrossRef Volltext
Downar, J., Crawley, AP, Mikulis, DJ und Davis, KD (2002). Ein kortikales Netzwerk, das für Stimulus Salience in einem neutralen Verhaltenskontext über mehrere sensorische Modalitäten hinweg empfindlich ist. In: J. Neurophysiol. 87, 615–620.
Pubmed Zusammenfassung / Pubmed Volltext
Drummond, L. und Shomstein, S. (2010). Objektbasierte Aufmerksamkeit: verschiebung oder Unsicherheit? Atten. Wahrnehmung. In: Psychophys. 72, 1743–1755.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
Egeth, H. E. und Yantis, S. (1997). Visuelle Aufmerksamkeit: Kontrolle, Repräsentation und Zeitverlauf. Jahr. In: Rev. Psychol. 48, 269–297.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
Eriksen, CW und Hoffman, JE (1972). Zeitliche und räumliche Merkmale der selektiven Codierung von visuellen Anzeigen. Wahrnehmung. In: Psychophys. 12, 201–204.Folk, C. L., Leber, A. B. und Egeth, H. E. (2002). Hat dich zum Blinzeln gebracht! Kontingente Aufmerksamkeitserfassung erzeugt ein räumliches Blinzeln. Wahrnehmung. In: Psychophys. 64, 741–753.
Pubmed Zusammenfassung / Pubmed Volltext
Folk, C. L., Remington, R. W. und Johnston, J. C. (1992). Unwillkürliche verdeckte Orientierung ist abhängig von Aufmerksamkeitssteuerungseinstellungen. In: J. Exp. Psychol. Summen. Wahrnehmung. Durchführen. 18, 1030–1044.
Pubmed Abstract / Pubmed Full Text
Friedrich, F. J., Egly, R., Rafal, R. D. und Beck, D. (1998). Räumliche Aufmerksamkeitsdefizite beim Menschen: Ein Vergleich von Läsionen des oberen parietalen und temporal-parietalen Übergangs. Neuropsychologie 12, 193-207.
Pubmed Zusammenfassung / Pubmed Volltext
Geng, JJ und Behrmann, M. (2002). Wahrscheinlichkeits-Cuing des Zielorts erleichtert die visuelle Suche implizit bei normalen Teilnehmern und Patienten mit hemispatialer Vernachlässigung. Psychol. sci. 13, 520–525.
Pubmed Zusammenfassung / Pubmed Volltext
Geng, JJ und Behrmann, M. (2005). Räumliche Wahrscheinlichkeit als Aufmerksamkeitshinweis bei der visuellen Suche. Wahrnehmung. In: Psychophys. 67, 1252–1268.
Pubmed Zusammenfassung / Pubmed Volltext
Geng, J. J. und Mangun, G. R. (2011). Die Aktivierung des rechten temporoparietalen Übergangs durch einen hervorstechenden kontextuellen Hinweis erleichtert die Zieldiskriminierung. Neuroimage 54, 594-601.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
Gibson, BS und Kelsey, EM (1998). Die reizgesteuerte Aufmerksamkeitserfassung hängt vom Aufmerksamkeitssatz für displayweite visuelle Merkmale ab. In: J. Exp. Psychol. Summen. Wahrnehmung. Durchführen. 24, 699–706.
Pubmed Abstract / Pubmed Full Text
Giesbrecht, B., Woldorff, M. G., Song, A. W. und Mangun, G. R. (2003). Neuronale Mechanismen der Top-Down-Kontrolle während der räumlichen und funktionalen Aufmerksamkeit. Neuroimage 19, 496-512.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
Gnadt, JW und Andersen, RA (1988). Gedächtnisbezogene motorische Planungsaktivität im posterioren parietalen Kortex von Makaken. Verwendbar bis. Brain Res. 70, 216-220.
Pubmed Zusammenfassung / Pubmed Volltext
Gottlieb, J. und Balan, P. (2010). Aufmerksamkeit als Entscheidung im Informationsraum. Trends Cogn. Sci. 14, 240–248.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
Greenberg, AS, Esterman, M., Wilson, D., Serences, JT und Yantis, S. (2010). Kontrolle der räumlichen und merkmalsbasierten Aufmerksamkeit im frontoparietalen Kortex. In: J. Neurosci. 30, 14330–14339.
Pubmed Abstract | Pubmed Full Text / CrossRef Full Text
Hopfinger, J. B., Buonocore, M. H. und Mangun, G. R. (2000). Die neuronalen Mechanismen der Top-Down-Aufmerksamkeitskontrolle. Nat. Neurowissenschaften. 3, 284–291.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
Ipata, A. E., Gee, A. L., Gottlieb, J., Bisley, J. W. und Goldberg, ME (2006). Lippenreaktionen auf einen Popout-Stimulus werden reduziert, wenn er offen ignoriert wird. Nat. Neurowissenschaften. 9, 1071–1076.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
Johnston, W. A. und Dark, V. J. (1986). Selektive Aufmerksamkeit. Jahr. In: Rev. Psychol. 37, 43–75.
Juola, JF, Koshino, H. und Warner, CB (1995). Kompromisse zwischen Aufmerksamkeitseffekten räumlicher Hinweise und abrupten Einsätzen. Wahrnehmung. In: Psychophys. 57, 333–342.
Pubmed Zusammenfassung / Pubmed Volltext
Kastner, S., Pinsk, MA, De Weerd, P., Desimone, R. und Ungerleider, LG (1999). Erhöhte Aktivität im menschlichen visuellen Kortex während der gerichteten Aufmerksamkeit in Abwesenheit von visueller Stimulation. Neuron 22, 751-761.
Pubmed Zusammenfassung / Pubmed Volltext / CrossRef Volltext
Koechlin, E., Ody, C. und Kouneiher, F. (2003). Die Architektur der kognitiven Kontrolle im menschlichen präfrontalen Kortex. Wissenschaft 302, 1181-1185.
Pubmed Zusammenfassung / Pubmed Volltext / CrossRef Volltext
Koshino, H., Warner, CB und Juola, JF (1992). Relative Wirksamkeit von zentralen, peripheren und abrupt einsetzenden Signalen in der visuellen Aufmerksamkeit. In: Q. J. Exp. Psychol. A 45, 609-631.
Pubmed Zusammenfassung / Pubmed Volltext
Liu, T., Slotnick, SD, Serences, JT und Yantis, S. (2003). Kortikale Mechanismen der merkmalsbasierten Aufmerksamkeitskontrolle. In: Cereb. Kapitel 13, 1334-1343.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Marois, R., Leung, H. C. und Gore, J. C. (2000). Ein reizgesteuerter Ansatz zur Objektidentitäts- und Ortsverarbeitung im menschlichen Gehirn. Neuron 25, 717-728.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
McFire, J. und Zangwill, O. L. (1960). Visuell-konstruktive Behinderungen im Zusammenhang mit Läsionen der linken Gehirnhälfte. Gehirn 82, 243-260.
Moore, C. M. und Egeth, H. (1998). Wie wirkt sich merkmalsbasierte Aufmerksamkeit auf die visuelle Verarbeitung aus? In: J. Exp. Psychol. Summen. Wahrnehmung. Durchführen. 24, 1296–1310.
Pubmed Zusammenfassung / Pubmed Volltext
Moran, J., und Desimone, R. (1985). Selektive Aufmerksamkeit und visuelle Verarbeitung im extrastriaten Kortex. Wissenschaft 229, 782-784.
Pubmed Zusammenfassung / Pubmed Volltext / CrossRef Volltext
O’Craven, K., Rosen, BR, Kwong, KK, Treisman, A. und Savoy, RL (1997). Freiwillige Aufmerksamkeit moduliert die fMRT-Aktivität im menschlichen MT-MST. Neuron 18, 591-598.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
Piercy, M. (1964). Die Auswirkungen zerebraler Läsionen auf die intellektuelle Funktion: ein Überblick über aktuelle Forschungstrends. Br. J. Psychiatrie 110, 310-352.
Pubmed Zusammenfassung / Pubmed Volltext
Posner, M. I. (1980). Orientierung der Aufmerksamkeit. In: Q. J. Exp. Psychol. 32, 3–25.
Pubmed Abstract / Pubmed Full Text
Posner, M. I., Snyder, C. R. und Davidson, B. J. (1980). Aufmerksamkeit und die Erkennung von Signalen. In: J. Exp. Psychol. 109, 160–174.
Pubmed Abstract / Pubmed Full Text
Posner, M. I., Walker, J. A., Friedrich, F. J. und Rafal, R. D. (1984). Auswirkungen von Parietalverletzungen auf die verdeckte Ausrichtung der Aufmerksamkeit. In: J. Neurosci. 4, 1863–1874.
Pubmed Zusammenfassung / Pubmed Volltext
Serences, J. T., Shomstein, S., Leber, A. B., Golay, X., Egeth, H. E. und Yantis, S. (2005). Koordination der freiwilligen und reizgesteuerten Aufmerksamkeitskontrolle im menschlichen Kortex. Psychol. Sci. 16, 114–122.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
Shaw, ML (1978). Ein Kapazitätszuordnungsmodell für die Reaktionszeit. In: J. Exp. Psychol. Summen. Wahrnehmung. Durchführen. 4, 586–598.
Shomstein, S. und Behrmann, M. (2006). Kortikale Systeme, die visuelle Aufmerksamkeit sowohl für Objekte als auch für räumliche Orte vermitteln. Prok. Natl. Acad. Sci. U.S.A. 103, 11387-11392.
Pubmed Zusammenfassung / Pubmed Volltext / CrossRef Volltext
Shomstein, S., Lee, J. und Behrmann, M. (2010). Aufmerksamkeitsführung von oben nach unten und von unten nach oben: Untersuchung der Rolle der dorsalen und ventralen parietalen Kortizes. Verwendbar bis. Brain Res. 206, 197-208.
Pubmed Zusammenfassung / Pubmed Volltext / CrossRef Volltext
Shomstein, S. und Yantis, S. (2004a). Konfigurative und kontextuelle Priorisierung in der objektbasierten Aufmerksamkeit. Psychon. Bull. Offenbarung 11, 247-253.
Pubmed Zusammenfassung / Pubmed Volltext
Shomstein, S. und Yantis, S. (2004b). Die Kontrolle der Aufmerksamkeit verschiebt sich im menschlichen Kortex zwischen Sehen und Hören. In: J. Neurosci. 24, 10702–10706.
Pubmed Zusammenfassung / Pubmed Volltext / CrossRef Volltext
Shomstein, S. und Yantis, S. (2006). Der parietale Kortex vermittelt die freiwillige Kontrolle der räumlichen und nichträumlichen auditorischen Aufmerksamkeit. In: J. Neurosci. 26, 435–439.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
Silver, M. A., Ress, D., und Heeger, D. J. (2005). Topographische Karten der visuellen räumlichen Aufmerksamkeit im menschlichen parietalen Kortex. In: J. Neurophysiol. 94, 1358–1371.
Pubmed Abstract | Pubmed Full Text / CrossRef Full Text
Tamber-Rosenau, BJ, Esterman, M., Chiu, Y.-C. und Yantis, S. (2011). Kortikale Mechanismen der kognitiven Kontrolle zur Verschiebung der Aufmerksamkeit im Seh- und Arbeitsgedächtnis. J. Cogn. Neurowissenschaften. 23, 2905–2919.
Pubmed Zusammenfassung | Pubmed Volltext / CrossRef Volltext
Theeuwes, J. (1991). Exogene und endogene Kontrolle der Aufmerksamkeit: die Wirkung von visuellen Ein- und Ausgängen. Wahrnehmung. In: Psychophys. 49, 83–90.
Pubmed Zusammenfassung / Pubmed Volltext
Vallar, G. und Perani, D. (1986). Die Anatomie der einseitigen Vernachlässigung nach Schlaganfallläsionen der rechten Hemisphäre. Eine klinische / CT-Scan-Korrelationsstudie am Menschen. Neuropsychologie 24, 609-622.
Pubmed Zusammenfassung / Pubmed Volltext / CrossRef Volltext
Wojciulik, E. und Kanwisher, N. (1999). Die Allgemeinheit der parietalen Beteiligung an der visuellen Aufmerksamkeit. Neuron 23, 747-764.
Pubmed Zusammenfassung / Pubmed Volltext / CrossRef Volltext
Yantis, S. und Jonides, J. (1984). Abrupte visuelle Onsets und selektive Aufmerksamkeit: Beweise aus der visuellen Suche. In: J. Exp. Psychol. Summen. Wahrnehmung. Durchführen. 10, 601–621.
Pubmed Zusammenfassung / Pubmed Volltext
Yantis, S. und Jonides, J. (1990). Abrupte visuelle Einbrüche und selektive Aufmerksamkeit: freiwillige versus automatische Zuordnung. In: J. Exp. Psychol. Summen. Wahrnehmung. Durchführen. 16, 121–134.
Pubmed Zusammenfassung / Pubmed Volltext
Yantis, S., Schwarzbach, J., Serences, J. T., Carlson, RL, Steinmetz, MA, Pekar, JJ und Courtney, SM (2002). Transiente neuronale Aktivität im menschlichen parietalen Kortex während räumlicher Aufmerksamkeitsverschiebungen. Nat. Neurowissenschaften. 5, 995–1002.
Pubmed Zusammenfassung / Pubmed Volltext / CrossRef Volltext
Yantis, S. und Serences, JT (2003). Kortikale Mechanismen der raum- und objektbasierten Aufmerksamkeitskontrolle. Curr. Opin. Neurobiol. 13, 187–193.
Pubmed Zusammenfassung / Pubmed Volltext / CrossRef Volltext