oppimistavoitteet
tämän osion loppuun mennessä voit:
- selitä glykolyysin prosessit
- kuvaa pyruvaattimolekyylin reitti Krebs-syklin läpi
- selitä elektronien kuljetus elektroninsiirtoketjun kautta
- kuvaa ATP: n tuotantoprosessia oksidatiivisen fosforylaation kautta
- Yhteenveto glukoneogeneesin prosessista
hiilihydraatit ovat hiili -, vety-ja happiatomeista koostuvia orgaanisia molekyylejä. Hiilihydraattiperheeseen kuuluu sekä yksinkertaisia että monimutkaisia sokereita. Glukoosi ja fruktoosi ovat esimerkkejä yksinkertaisista sokereista, ja tärkkelys, glykogeeni ja selluloosa ovat kaikki esimerkkejä monimutkaisista sokereista. Kompleksisokereita kutsutaan myös polysakkarideiksi ja ne koostuvat useista monosakkaridimolekyyleistä. Polysakkaridit toimivat energian varastointina (esim.tärkkelys ja glykogeeni) ja rakenneosina (esim. kitiini hyönteisissä ja selluloosa kasveissa).
ruoansulatuksen aikana hiilihydraatit pilkkoutuvat yksinkertaisiksi, liukoisiksi sokereiksi, jotka voivat kulkeutua suolen seinämän läpi verenkiertoelimistöön kuljetettavaksi koko kehoon. Hiilihydraattien ruoansulatus alkaa suussa syljen amylaasin vaikutuksesta tärkkelykseen ja päättyy monosakkaridien imeytymiseen ohutsuolen epiteelin poikki. Kun imeytyneet monosakkaridit kuljetetaan kudoksiin, soluhengitysprosessi alkaa (kuva 1). Tässä jaksossa keskitytään ensin glykolyysiin eli prosessiin, jossa monosakkaridi glukoosi hapettuu, jolloin sen sidoksiin varastoitunut energia vapautuu ATP: n tuottamiseksi.
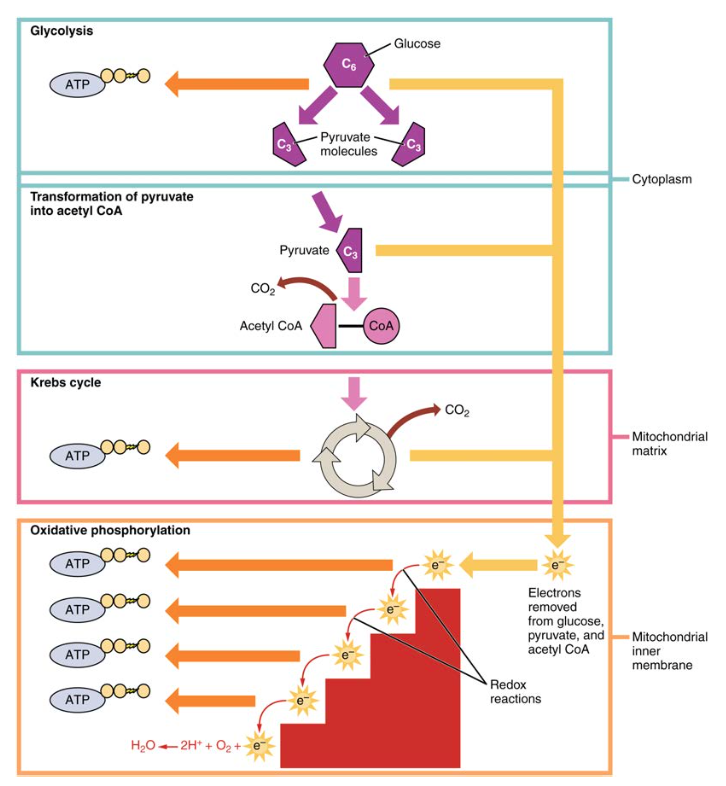
kuva 1. Soluhengitys hapettaa glukoosimolekyylit glykolyysin, Krebs-kierron ja oksidatiivisen fosforylaation avulla, jolloin muodostuu ATP: tä.
glykolyysi
glukoosi on elimistön helposti saatavilla oleva energianlähde. Kun ruoansulatus hajottaa polysakkaridit alas monosakkarideiksi, mukaan lukien glukoosi, monosakkaridit kuljetetaan ohutsuolen seinämän yli verenkiertoelimistöön, joka kuljettaa ne maksaan. Maksassa maksasolut joko siirtävät glukoosin verenkiertoelimistön kautta tai varastoivat ylimääräisen glukoosin glykogeeniksi. Elimistön solut imevät verenkierrossa olevaa glukoosia insuliinivasteena ja glykolyysiksi kutsutun reaktiosarjan kautta siirtävät osan glukoosin energiasta ADP: hen, jolloin muodostuu ATP: tä (kuva 2). Glykolyysin viimeisessä vaiheessa syntyy tuote pyruvaatti.
glykolyysi alkaa glukoosin fosforyloinnilla heksokinaasin avulla muodostaen glukoosi-6-fosfaattia. Tässä vaiheessa käytetään yhtä ATP: tä, joka on fosfaattiryhmän luovuttaja. Fosfofruktokinaasin vaikutuksesta glukoosi-6-fosfaatti muuttuu fruktoosi-6-fosfaatiksi. Tässä vaiheessa toinen ATP luovuttaa fosfaattiryhmänsä, jolloin muodostuu fruktoosi-1,6-bisfosfaattia. Tämä kuusihiilinen sokeri pilkkoutuu muodostaen kaksi fosforyloitua kolmihiilistä molekyyliä, glyseraldehydi-3-fosfaatin ja dihydroksiasetonifosfaatin, jotka molemmat muuttuvat glyseraldehydi-3-fosfaatiksi. Glyseraldehydi-3-fosfaatti fosforyloidaan edelleen solussa olevan divetyfosfaatin luovuttamilla ryhmillä, jolloin muodostuu kolmihiilinen molekyyli 1,3-bisfosfoglyseraatti. Tämän reaktion energia tulee glyseraldehydi-3-fosfaatin hapettumisesta (elektronien poistamisesta). Pyruvaattiin johtavissa reaktioissa kaksi fosfaattiryhmää siirretään sitten kahteen ADPs: ään muodostaen kaksi ATPs: ää. Glykolyysissä käytetään siis kahta ATP: tä, mutta syntyy neljä ATP: tä, jolloin saadaan nettovoitto kahdesta ATP: stä ja kahdesta pyruvaattimolekyylistä. Hapen läsnä ollessa pyruvaatti jatkuu Krebs-sykliin (jota kutsutaan myös sitruunahappokierroksi tai trikarboksyylihappokierroksi (TCA), jossa lisäenergia erotetaan ja siirretään eteenpäin.
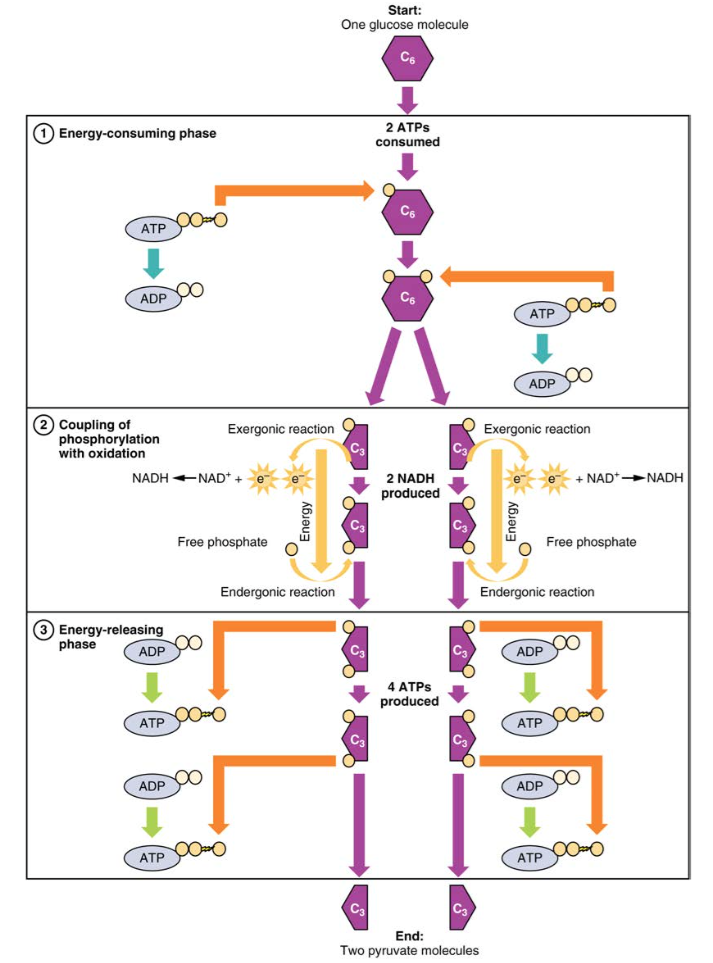
kuva 2. Glykolyysin energiaa kuluttavassa vaiheessa kuluu kaksi ATP: tä, jolloin glukoosimolekyylille siirtyy kaksi fosfaattia. Tämän jälkeen glukoosimolekyyli jakautuu kahdeksi kolmihiiliseksi yhdisteeksi, joista jokainen sisältää fosfaatin. Toisen vaiheen aikana jokaiseen kolmihiiliseen yhdisteeseen lisätään ylimääräinen fosfaatti. Energia tälle endergoniselle reaktiolle saadaan poistamalla (hapettamalla) kaksi elektronia kustakin kolmen hiilen yhdisteestä. Energiaa vapauttavassa vaiheessa fosfaatit poistetaan molemmista kolmihiilisistä yhdisteistä ja niitä käytetään neljän ATP-molekyylin tuottamiseen.
katso videolta tietoa glykolyysistä:
glykolyysi voidaan jakaa kahteen vaiheeseen: energian kuluttamiseen (jota kutsutaan myös kemialliseksi pohjustukseksi) ja energian tuottamiseen. Ensimmäinen vaihe on energiaa kuluttava vaihe, joten reaktion käynnistämiseen tarvitaan kaksi ATP-molekyyliä jokaista glukoosimolekyyliä kohti. Reaktion lopussa syntyy kuitenkin neljä ATP-Energiamolekyyliä, jolloin syntyy kahden ATP-energiamolekyylin nettovoitto.
glykolyysi voidaan ilmaista seuraavalla yhtälöllä:
\text{Glucose}+2\text{ATP}+2\text{NAD}^{+}+4\text{ADP}+2\text{P}_{i}\to\text{Pyruvate}+4\text{ATP}+2\text{NADH}+2\text{H}^{+}
tämän yhtälön mukaan glukoosi hajoaa yhdessä ATP: n (energianlähde), NAD+: n (elektronisenseptorina toimiva koentsyymi) ja epäorgaanisen fosfaatin kanssa kahdeksi pyruvaattimolekyyliksi, jolloin syntyy neljä ATP—molekyyliä—kahden ATP: n nettotuotolla-ja kaksi energiaa sisältävää NADH-koentsyymiä. Tässä prosessissa syntyvää NADH: ta käytetään myöhemmin ATP: n tuottamiseen mitokondrioissa. Tärkeää on, että tämän prosessin loppuun mennessä yksi glukoosimolekyyli tuottaa kaksi pyruvaattimolekyyliä, kaksi korkeaenergistä ATP-molekyyliä ja kaksi elektronia kuljettavaa NADH-molekyyliä.
seuraavat glykolyysiä koskevat keskustelut sisältävät reaktioista vastaavat entsyymit. Kun glukoosi siirtyy soluun, maksassa oleva heksokinaasientsyymi (tai glukokinaasi) lisää nopeasti fosfaattia muuttaakseen sen glukoosi-6-fosfaatiksi. Kinaasi on entsyymityyppi, joka lisää substraattiin fosfaattimolekyylin (tässä tapauksessa glukoosin, mutta se voi olla totta myös muille molekyyleille). Tämä muunnosvaihe vaatii yhden ATP: n ja lähinnä vangitsee glukoosin soluun, estäen sen kulkeutumisen takaisin plasmakalvon läpi, jolloin glykolyysi voi edetä. Se toimii myös pitoisuuden gradientin ylläpitämiseksi veren korkeammalla glukoosipitoisuudella kuin kudoksissa. Määrittämällä tämän pitoisuusgradientin veren glukoosi pystyy virtaamaan korkean pitoisuuden alueelta (veri) alhaisen pitoisuuden alueelle (kudokset) joko käytettäväksi tai varastoitavaksi. Heksokinaasia esiintyy lähes jokaisessa kehon kudoksessa. Glukokinaasi taas ilmenee kudoksissa, jotka ovat aktiivisia veren glukoosipitoisuuden ollessa korkea, kuten maksassa. Heksokinaasilla on suurempi affiniteetti glukoosiin kuin glukokinaasilla, minkä vuoksi se kykenee muuntamaan glukoosia nopeammin kuin glukokinaasi. Tämä on tärkeää, kun glukoosipitoisuus on hyvin alhainen elimistössä, koska se mahdollistaa glukoosin matkustaa mieluiten niihin kudoksiin, jotka tarvitsevat sitä enemmän.
glykolyysin ensimmäisen vaiheen seuraavassa vaiheessa glukoosi-6-fosfaatti-isomeraasientsyymi muuttaa glukoosi-6-fosfaatin fruktoosi-6-fosfaatiksi. Glukoosin tavoin myös fruktoosi on kuutta hiiltä sisältävä sokeri. Fosfofruktokinaasi-1-entsyymi lisää sitten vielä yhden fosfaatin muuttaakseen fruktoosi-6-fosfaatin fruktoosiksi-1-6-bisfosfaatti, toinen kuusihiilinen sokeri, käyttäen toista ATP-molekyyliä. Aldolaasi hajottaa tämän fruktoosin-1-6-bisfosfaatti kahdeksi kolmihiiliseksi molekyyliksi, glyseraldehydi-3-fosfaatiksi ja dihydroksiasetonifosfaatiksi. Tämän jälkeen triosefosfaatti-isomeraasientsyymi muuntaa dihydroksiasetonifosfaatin toiseksi glyseraldehydi-3-fosfaattimolekyyliksi. Siksi tämän kemiallisesti pohjustavan eli energiaa kuluttavan vaiheen loppuun mennessä yksi glukoosimolekyyli hajoaa kahdeksi glyseraldehydi-3-fosfaattimolekyyliksi.
glykolyysin toinen vaihe, energiaa tuottava vaihe, luo glykolyysin tuottaman energian. Glyseraldehydi-3-fosfaattidehydrogenaasi muuntaa jokaisen
energiaa kuluttavan vaiheen aikana syntyneen kolmihiilisen glyseraldehydi-3-fosfaatin 1,3-bisfosfoglyseraatiksi. Tässä reaktiossa vapautuu elektroni, jonka NAD+ poimii talteen, jolloin syntyy NADH-molekyyli. NADH on ATP: n tavoin suurenerginen molekyyli, mutta toisin kuin ATP, solu ei käytä sitä energiavaluuttana. Koska glyseraldehydi-3-fosfaattimolekyylejä on kaksi, tässä vaiheessa syntetisoidaan kaksi NADH-molekyyliä. Jokainen 1,3-bisfosfoglyseraatti defosforyloidaan (eli fosfaatti poistetaan) fosfoglyseraattikinaasin vaikutuksesta 3-fosfoglyseraatiksi. Jokainen tässä reaktiossa vapautuva fosfaatti voi muuntaa yhden ADP-molekyylin yhdeksi korkeaenergiseksi ATP-molekyyliksi, jolloin saadaan kaksi ATP-molekyyliä.
fosfoglyseraattimutaasientsyymi muuttaa tämän jälkeen 3-fosfoglyseraattimolekyylit 2-fosfoglyseraatiksi. Tämän jälkeen enolaasientsyymi vaikuttaa 2-fosfoglyseraattimolekyyleihin muuttaen ne fosfoenolipyruvaattimolekyyleiksi. Glykolyysin viimeisessä vaiheessa kaksi fosfoenolipyruvaattimolekyyliä defosforyloidaan pyruvaattikinaasin avulla, jolloin syntyy kaksi pyruvaattimolekyyliä ja kaksi ATP-molekyyliä.
yhteenvetona voidaan todeta, että yksi glukoosimolekyyli hajoaa kahdeksi pyruvaattimolekyyliksi ja muodostaa glykolyysin avulla kaksi nettoatp-molekyyliä ja kaksi NADH-molekyyliä. Siksi glykolyysi tuottaa solulle energiaa ja luo pyruvaattimolekyylejä, joita voidaan jalostaa edelleen aerobisen Krebs-syklin (jota kutsutaan myös sitruunahappokierroksi tai trikarboksyylihappokierroksi), muuttaa maitohapoksi tai alkoholiksi (hiivassa) käymisen avulla tai käyttää myöhemmin glukoosin synteesissä glukoneogeneesin avulla.
anaerobinen hengitys
kun happea on vähän tai sitä ei ole, pyruvaatti siirtyy anaerobiseen kulkureittiin. Näissä reaktioissa pyruvaatti voi muuttua maitohapoksi. Sen lisäksi, että tämä reitti tuottaa ylimääräistä ATP: tä, se auttaa pitämään pyruvaattipitoisuuden alhaisena, joten glykolyysi jatkuu ja se hapettaa NADH: n glykolyysin tarvitsemaksi NAD+: ksi. Tässä reaktiossa maitohappo korvaa hapen lopullisena elektronisenseptorina. Anaerobista hengitystä tapahtuu useimmissa elimistön soluissa, kun happea on vähän tai mitokondriot puuttuvat tai eivät toimi. Esimerkiksi koska punasoluilla (punasoluilla) ei ole mitokondrioita, niiden on tuotettava ATP: tä anaerobisesta hengityksestä. Tämä on tehokas polku ATP tuotantoa lyhyitä aikoja, jotka vaihtelevat sekunneista muutamaan minuuttiin. Tuotettu maitohappo diffundoituu plasmaan ja kulkeutuu maksaan, jossa se Cori-kierron kautta muuttuu takaisin pyruvaatiksi tai glukoosiksi. Samoin kun henkilö harjoittaa, lihakset käyttävät ATP nopeammin kuin happea voidaan toimittaa heille. Ne ovat riippuvaisia glykolyysistä ja maitohapon tuotannosta nopeaan ATP: n tuotantoon.
aerobinen hengitys
hapen läsnä ollessa pyruvaatti voi päästä Krebs-sykliin, jossa saadaan lisäenergiaa elektronien siirtyessä pyruvaatista reseptoreihin NAD+, GDP ja FAD hiilidioksidin ollessa ”jätetuote” (kuva 3). NADH ja FADH2 siirtävät elektroneja elektroninsiirtoketjuun, joka käyttää siirretyn energian ATP: n tuottamiseen. Elektroninsiirtoketjun terminaalivaiheena happi toimii terminaalielektronien hyväksyjänä ja luo mitokondrioiden sisälle vettä.
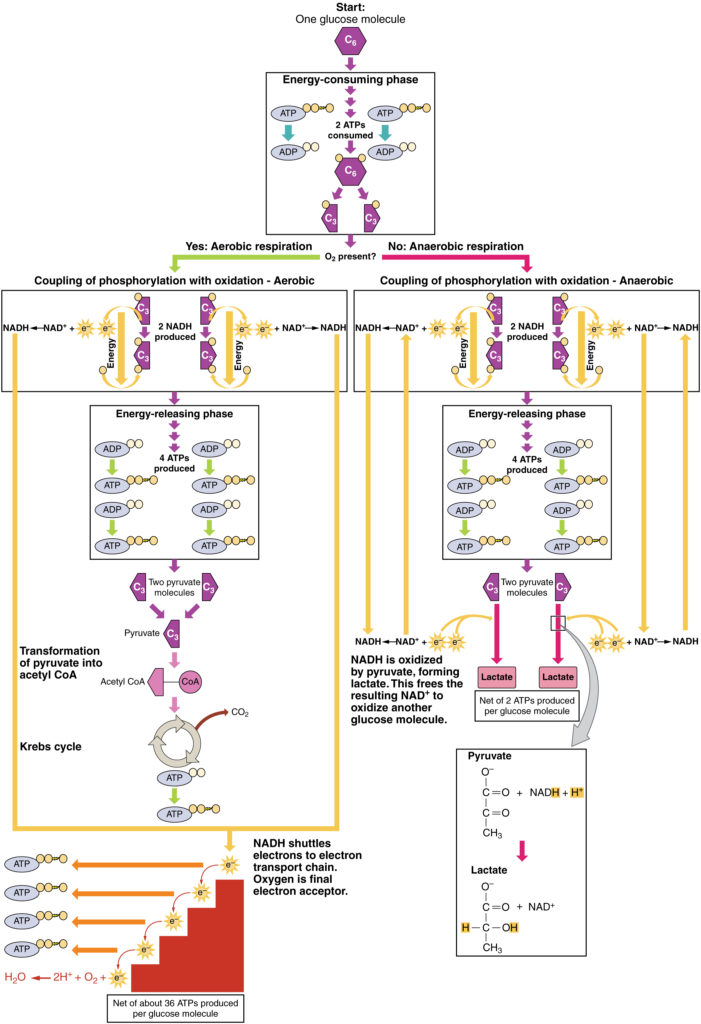
kuva 3. Klikkaa nähdäksesi suuremman kuvan. Anaerobisen hengityksen prosessi muuntaa glukoosin kahdeksi laktaattimolekyyliksi hapen puuttuessa tai punasolujen sisällä, joilta puuttuu mitokondriot. Aerobisen hengityksen aikana glukoosi hapetetaan kahdeksi pyruvaattimolekyyliksi.
Krebs-sykli/Sitruunahapposykli/Trikarboksyylihapposykli
glykolyysin aikana syntyvät pyruvaattimolekyylit kulkeutuvat mitokondrion kalvon läpi sisempään mitokondriomatriisiin, jossa entsyymit metaboloivat ne Krebs-sykliksi kutsuttua reittiä (Kuva 4). Krebsin sykliä kutsutaan yleisesti myös sitruunahapposykliksi tai trikarboksyylihapposykliksi (TCA). Krebs-syklissä syntyy suurienergiaisia molekyylejä, kuten ATP, NADH ja FADH2. Tämän jälkeen NADH ja FADH2 siirtävät mitokondrioissa elektroneja elektroninsiirtoketjun läpi tuottaakseen lisää ATP-molekyylejä.
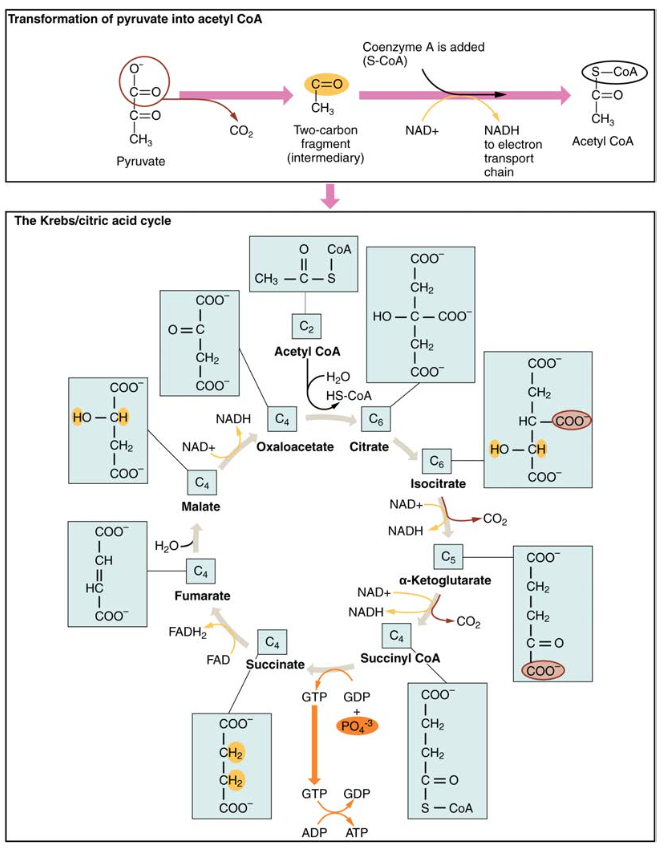
kuva 4. Krebs-syklissä jokainen glykolyysissä syntyvä pyruvaatti muuttuu kaksihiiliseksi asetyyli-CoA-molekyyliksi. Asetyyli-CoA käsitellään järjestelmällisesti läpi syklin ja tuottaa suurienergisiä NADH -, FADH2-ja ATP-molekyylejä.
Katso tästä animaatiosta Krebsin sykli.
glykolyysissä syntyvä kolmihiilinen pyruvaattimolekyyli siirtyy sytoplasmasta mitokondriomatriisiin, jossa pyruvaattidehydrogenaasientsyymi muuntaa sen kaksihiiliseksi asetyylikoentsyymi-A: ksi (asetyyli-CoA) molekyyliksi. Reaktio on oksidatiivinen dekarboksylaatioreaktio. Se muuntaa kolmihiilisen pyruvaatin kaksihiiliseksi asetyyli-CoA-molekyyliksi vapauttaen hiilidioksidia ja siirtäen kaksi elektronia, jotka yhdistyvät NAD+: n kanssa muodostaen NADH: ta. Asetyyli-CoA siirtyy Krebs-sykliin yhdistymällä nelihiilisen molekyylin, oksaloasetaatin, kanssa muodostaen kuusihiilisen molekyylisitraatin eli sitruunahapon, jolloin samalla vapautuu koentsyymi-A-molekyyli.
kuusihiilisitraattimolekyyli muuttuu systemaattisesti ensin viisihiiliseksi molekyyliksi ja sitten nelihiiliseksi molekyyliksi päättyen oksaloasetaattiin, joka on syklin alku. Matkan varrella kukin sitraattimolekyyli tuottaa yhden ATP: n, yhden FADH2: n ja kolme NADH: ta. Fadh2 ja NADH siirtyvät hapettavaan fosforylaatiojärjestelmään, joka sijaitsee sisemmässä mitokondrioiden kalvossa. Lisäksi Krebs-sykli toimittaa lähtöaineet proteiinien ja rasvojen prosessointiin ja hajottamiseen.
Krebs-syklin käynnistämiseksi sitraattisyntaasi yhdistää asetyyli-CoA: n ja oksaloasetaatin muodostaen kuusihiilisen sitraattimolekyylin; CoA vapautuu myöhemmin ja voi yhdistyä toisen pyruvaattimolekyylin kanssa aloittaa syklin uudelleen. Akonitaasientsyymi muuttaa sitraatin isositraatiksi. Oksidatiivisen dekarboksylaation kahdessa peräkkäisessä vaiheessa muodostuu kaksi CO2-molekyyliä ja kaksi NADH-molekyyliä, kun isositraattidehydrogenaasi muuntaa isositraatin viisihiiliseksi α-ketoglutaraattidehydrogenaasin katalysoimana ja muuttaa sen nelihiiliseksi sukkinyyli-CoA: ksi α-ketoglutaraattidehydrogenaasin vaikutuksesta. Tämän jälkeen sukkinyyli-CoA-dehydrogenaasientsyymi muuntaa sukkinyyli-CoA: n sukkinaatiksi ja muodostaa korkeaenergisen molekyylin GTP: n, joka siirtää energiansa ADP: hen tuottaakseen ATP: tä. Tämän jälkeen sukkinaattidehydrogenaasi muuntaa sukkinaatin fumaraatiksi, jolloin muodostuu fadh2-molekyyli. Tämän jälkeen fumaraasi muuntaa fumaraatin malaatiksi, jonka malaattidehydrogenaasi muuntaa takaisin oksaloasetaatiksi pelkistäen samalla NAD+: n NADH: ksi. Oksaloasetaatti on valmis yhdistymään seuraavan asetyyli-CoA: n kanssa Krebs-syklin aloittamiseksi uudelleen (KS.Kuva 4). Jokaista kierrosta kohti syntyy kolme NADH: ta, yksi ATP (GTP: n kautta) ja yksi FADH2. Jokainen pyruvaatin hiili muuttuu CO2: ksi, joka vapautuu hapettavan (aerobisen) hengityksen sivutuotteena.
oksidatiivinen fosforylaatio ja Elektroninsiirtoketju
elektroninsiirtoketju (ETC) käyttää Krebs-syklin tuottamaa NADH: ta ja FADH2: ta ATP: n tuottamiseen. NADH: n ja FADH2: n elektronit siirtyvät entsymaattisten reaktioiden sarjassa mitokondrion sisempään kalvoon upotettujen proteiinikompleksien kautta. Elektroninsiirtoketju koostuu neljästä entsyymikompleksista (kompleksi I – kompleksi IV) ja kahdesta koentsyymistä (ubikinoni ja sytokromi c), jotka toimivat elektroninkantajina ja protonipumppuina, joita käytetään siirtämään h+ – ioneja sisä-ja ulompien mitokondrioiden väliseen tilaan (kuva 5). ETC yhdistää elektronien siirron luovuttajan (kuten NADH) ja elektronisenseptorin (kuten O2) välillä protonien (H+ ionien) siirtoon sisemmän mitokondriokalvon poikki mahdollistaen hapettavan fosforylaation prosessin. Hapen läsnä ollessa energiaa johdetaan vaiheittain elektroninkantajien läpi keräämään vähitellen tarvittava energia fosfaatin liittämiseksi ADP: hen ja ATP: n tuottamiseksi. Molekyylihapen eli O2: n rooli on ETC: n terminaalielektroniakseptorina. Tämä tarkoittaa sitä, että kun elektronit ovat kulkeneet koko ETC: n läpi, ne on siirrettävä toiselle, erilliselle molekyylille. Nämä matriisin elektronit, O2-ja H+ – ionit yhdistyvät muodostaen uusia vesimolekyylejä. Tämä on perusta tarpeellesi hengittää happea. Ilman happea elektronivirtaus ETC: n läpi lakkaa.
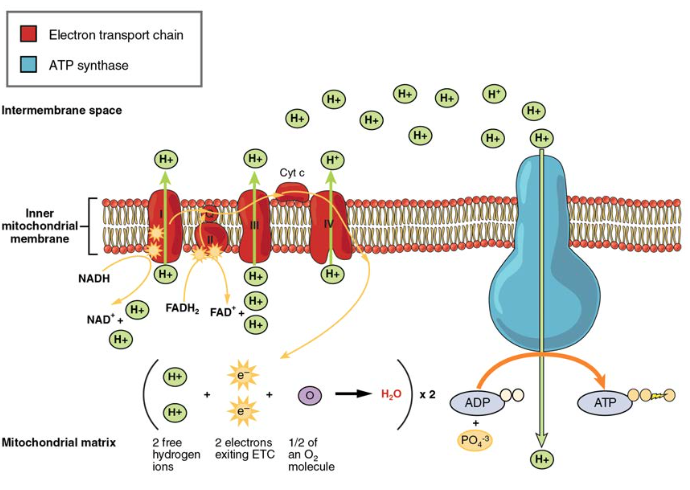
kuva 5. Elektroninsiirtoketju on sarja elektroninkantajia ja ionipumppuja, joiden avulla h+ – ioneja pumpataan ulos sisemmästä mitokondriomatriisista.
katso videolta tietoa elektroninsiirtoketjusta.
kunkin kantajan kautta kulkee NADH: sta ja FADH2: sta vapautuneet elektronit, jotka pelkistyvät vastaanottaessaan elektronin ja hapettuvat siirtäessään sen seuraavalle kantajalle. Jokainen näistä reaktioista vapauttaa pienen määrän
energiaa, jota käytetään H+ – ionien pumppaamiseen sisäkalvon poikki. Näiden protonien kertyminen kalvojen väliseen tilaan luo protonigradientin mitokondrion matriisiin nähden.
myös sisempään mitokondriokalvoon on upotettu hämmästyttävä proteiinihuokoskompleksi, jota kutsutaan ATP-syntaasiksi. Tehokkaasti se on turbiini, joka saa voimansa H+ – ionien virtauksesta sisäkalvon läpi alas gradienttia ja mitokondrioiden matriisiin. H + – ionien kulkiessa kompleksin halki kompleksin akseli pyörii. Tämä kierto mahdollistaa ATP-syntaasin muiden osien kannustamisen ADP: hen ja Pi: hen luomaan ATP: tä. Laskettaessa aerobisen hengityksen kautta tuotetun ATP: n kokonaismäärää glukoosimolekyyliä kohti on tärkeää muistaa seuraavat seikat:
- glykolyysin kautta syntyy kaksi ATP: tä (neljä tuotetaan ja kaksi kulutetaan energiaa kuluttavassa vaiheessa). Näitä kahta ATP: tä käytetään kuitenkin glykolyysin aikana syntyvän NADH: n kuljettamiseen sytoplasmasta mitokondrioihin. Siksi ATP: n nettotuotanto glykolyysin aikana on nolla.
- kaikissa glykolyysin jälkeisissä vaiheissa on tuotettujen ATP: n, NADH: n ja FADH2: n määrä kerrottava kahdella, jotta voidaan selvittää, miten kukin glukoosimolekyyli tuottaa kaksi pyruvaattimolekyyliä.
- ETC: ssä jokaista hapettunutta NADH: ta kohti tuotetaan noin kolme ATP: tä. Jokaista hapettunutta FADH2: ta kohti tuotetaan kuitenkin vain noin kaksi ATP: tä. Fadh2: n elektronit tuottavat vähemmän ATP: tä, koska ne alkavat ETC: n (kompleksi II) matalammasta pisteestä verrattuna NADH: n (kompleksi I) elektroneihin (kuva 5).
näin ollen jokaista aerobiseen hengitykseen tulevaa glukoosimolekyyliä kohti muodostuu yhteensä 36 ATP: tä (KS.kuva 6).
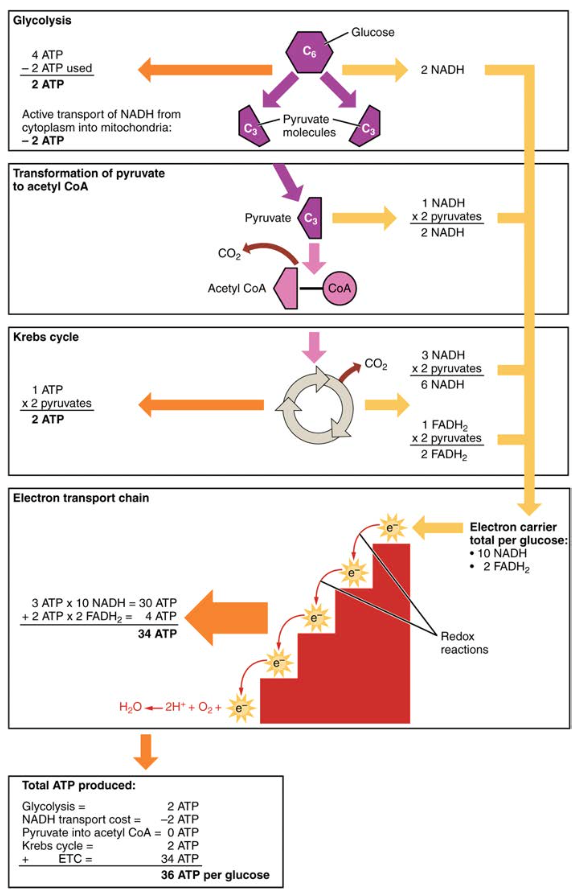
kuva 6. Hiilihydraattien aineenvaihduntaan liittyy glykolyysiä, Krebs-kiertoa ja elektroninsiirtoketjua.
glukoneogeneesi
glukoneogeneesi on uusien glukoosimolekyylien synteesi pyruvaatista, laktaatista, glyserolista tai alaniinin tai glutamiinin aminohapoista. Tämä prosessi tapahtuu pääasiassa maksassa alhaisen glukoosipitoisuuden aikana, toisin sanoen paaston, nälänhädän ja vähähiilihydraattisen ruokavalion olosuhteissa. Voidaan siis kysyä, miksi keho loisi jotain, minkä hajottamiseen se on juuri nähnyt melkoisesti vaivaa? Tietyt keskeiset elimet, mukaan lukien aivot, voivat käyttää vain glukoosia energianlähteenä; siksi on tärkeää, että elimistö ylläpitää veren glukoosipitoisuuden minimitasoa. Kun veren glukoosipitoisuus laskee alle tietyn pisteen, maksa syntetisoi uuden glukoosin nostaakseen veren pitoisuuden normaaliksi.
glukoneogeneesi ei ole pelkästään glykolyysin vastakohta. On joitakin merkittäviä eroja (kuva 7). Pyruvaatti on yleinen glukoneogeneesin lähtöaine. Ensin pyruvaatti muutetaan oksaloasetaatiksi. Oksaloasetaatti toimii tämän jälkeen substraattina fosfoenolipyruvaattikarboksikinaasientsyymille (PEPCK), joka muuttaa oksaloasetaatin fosfoenolipyruvaatiksi (Pep). Tästä vaiheesta glukoneogeneesi on lähes glykolyysin vastakohta. PEP muutetaan takaisin 2-fosfoglyseraatiksi, joka muuttuu 3-fosfoglyseraatiksi. Tämän jälkeen 3-fosfoglyseraatti muutetaan 1,3-bisfosfoglyseraatiksi ja sitten glyseraldehydi-3-fosfaatiksi. Kaksi glyseraldehydi-3-fosfaattimolekyyliä yhdistyy sitten fruktoosiksi-1-6-bisfosfaatti, joka muuttuu fruktoosi-6-fosfaatiksi ja sitten glukoosi-6-fosfaatiksi. Lopuksi sarja reaktioita tuottaa itse glukoosia. Glukoneogeneesissä (verrattuna glykolyysiin) heksokinaasientsyymi korvautuu glukoosi-6-fosfataasilla ja fosfofruktokinaasi-1-entsyymi fruktoosi-1,6-bisfosfataasilla. Tämä auttaa solua säätelemään glykolyysiä ja glukoneogeneesiä toisistaan riippumatta.
kuten lipolyysin yhteydessä käsitellään, rasvat voidaan hajottaa glyseroliksi, joka voidaan fosforyloida dihydroksiasetonifosfaatiksi tai DHAP: ksi. DHAP voi joko tulla glykolyyttiseen kulkureittiin tai sitä käytetään maksassa glukoneogeneesin substraattina.
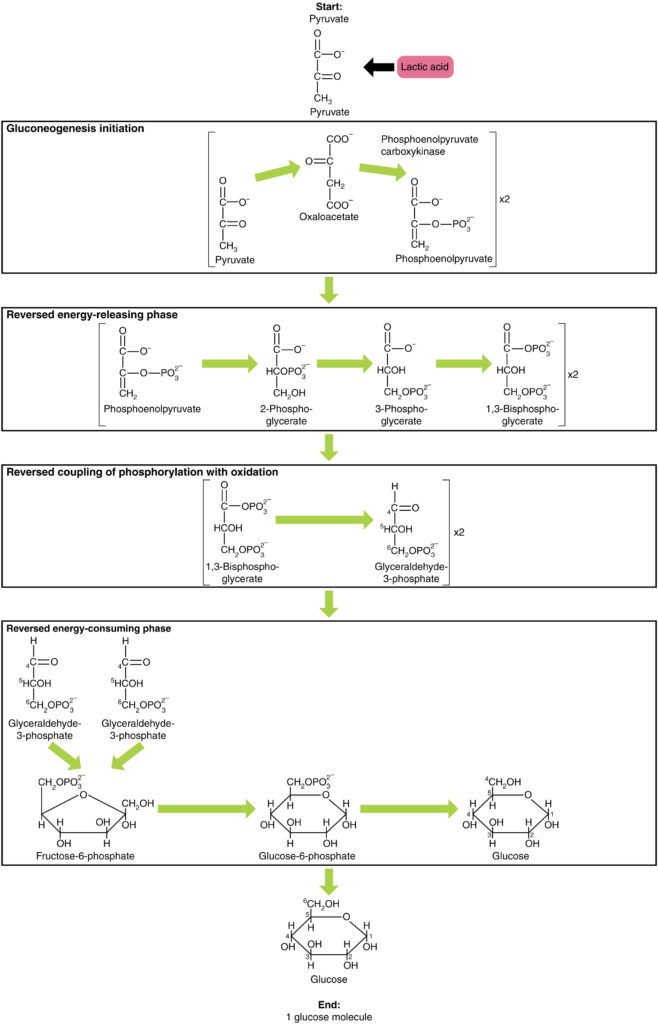
kuva 7. Klikkaa nähdäksesi suuremman kuvan. Glukoneogeneesi on glukoosin synteesi pyruvaatista, laktaatista, glyserolista, alaniinista tai glutamaatista.
ikääntyminen ja kehon aineenvaihdunta
ihmiskehon aineenvaihdunta hidastuu lähes 2 prosenttia vuosikymmenessä 30 ikävuoden jälkeen. Kehon koostumuksen muutokset, mukaan lukien vähärasvaisen lihasmassan väheneminen, ovat useimmiten vastuussa tästä vähenemisestä. Dramaattisin lihasmassan menetys ja sen seurauksena aineenvaihdunnan hidastuminen tapahtuu 50-70-vuotiaana. Lihasmassan menetys vastaa heikentynyttä voimaa, mikä on omiaan estämään senioreita harjoittamasta riittävää liikuntaa. Tämä johtaa positiivisen palautteen järjestelmään, jossa vähentynyt liikunta johtaa entistä enemmän lihasten menetykseen, mikä edelleen vähentää aineenvaihduntaa.
on useita asioita, joilla voidaan ehkäistä aineenvaihdunnan yleistä heikkenemistä ja taistella näiden heikkenemisen syklisyyttä vastaan. Näitä ovat syöminen aamiainen, syö pieniä aterioita usein, kuluttaa runsaasti vähärasvaista proteiinia, juomaveden pysyä sammutettua, käyttäessään (mukaan lukien voimaharjoittelu), ja saada tarpeeksi unta. Nämä toimenpiteet voivat auttaa pitämään energiatasot laskusta ja hillitä halu lisätä kalorien kulutusta liiallisesta snacking. Vaikka nämä strategiat eivät ole taattu ylläpitää aineenvaihduntaa, ne auttavat estämään lihasten menetys ja voivat lisätä energian tasoilla. Jotkut asiantuntijat ehdottavat myös sokerin välttämistä, mikä voi johtaa rasvan liikavarastoitumiseen. Myös mausteisista ruuista ja vihreästä teestä voi olla hyötyä. Koska stressi aktivoi kortisolin vapautumista ja kortisoli hidastaa aineenvaihduntaa, voi myös stressin välttäminen tai ainakin rentoutustekniikoiden harjoittaminen auttaa.
Lukukatsaus
Metaboliaentsyymit katalysoivat katabolisia reaktioita, jotka hajottavat ruoan sisältämiä hiilihydraatteja. Vapautuva energia käytetään virtaa soluja ja järjestelmiä, jotka muodostavat kehon. Ylimääräinen tai käyttämätön energia varastoidaan rasvaksi tai glykogeeniksi myöhempää käyttöä varten. Hiilihydraattien aineenvaihdunta alkaa suusta, jossa syljen amylaasientsyymi alkaa hajottaa kompleksisia sokereita monosakkarideiksi. Nämä voidaan sitten kuljettaa läpi suoliston kalvon verenkiertoon ja sitten kehon kudoksiin. Soluissa glukoosi, kuuden hiilen sokeri, prosessoidaan reaktiosarjan kautta pienemmiksi sokereiksi,ja molekyylin sisään varastoitunut energia vapautuu. Hiilihydraattikatabolian ensimmäinen vaihe on glykolyysi, joka tuottaa pyruvaattia, NADH: ta ja ATP: tä. Anaerobisissa olosuhteissa pyruvaatti voidaan muuttaa laktaatiksi glykolyysin pitämiseksi toiminnassa. Pyruvaatti siirtyy aerobisissa olosuhteissa Krebs-sykliin, jota kutsutaan myös sitruunahappokierroksi tai trikarboksyylihappokierroksi. ATP: n lisäksi Krebsin syklissä syntyy suurienergiaisia fadh2-ja NADH-molekyylejä, jotka luovuttavat elektroneja oksidatiiviseen fosforylaatioprosessiin, jossa syntyy enemmän suurienergiaisia ATP-molekyylejä. Jokaista glykolyysissä käsiteltävää glukoosimolekyyliä kohti voidaan aerobisen hengityksen avulla luoda 36 ATPs: n netto.
anaerobisissa olosuhteissa ATP: n tuotanto rajoittuu glykolyysissä syntyviin. Glykolyysissä syntyy yhteensä neljä ATP: tä, mutta glykolyysin aloittamiseen tarvitaan kaksi, joten kahden ATP-molekyylin nettosaanto on mahdollinen.
alhaisen glukoosin olosuhteissa, kuten paastossa, nälkiintymisessä tai vähähiilihydraattisessa ruokavaliossa, glukoosia voidaan syntetisoida laktaatista, pyruvaatista, glyserolista, alaniinista tai glutamaatista. Tämä prosessi, jota kutsutaan glukoneogeneesiksi, on lähes glykolyysin vastakohta ja palvelee glukoosimolekyylien muodostamista glukoosista riippuvaisiin elimiin, kuten aivoihin, kun glukoositaso laskee alle normaalin.
Self Check
vastaa alla oleviin kysymyksiin nähdäksesi, kuinka hyvin ymmärrät edellisessä jaksossa käsitellyt aiheet.
kriittiset Ajattelukysymykset
- selittävät, miten glukoosi metaboloituu ATP: n tuottamiseksi.
- Keskustele mekanismista, jolla solut luovat pitoisuusgradientin, joka varmistaa glukoosin jatkuvan imeytymisen verenkiertoon.
Sanasto
polysakkaridit: monista monosakkarideista koostuvat kompleksiset hiilihydraatit
monosakkaridi: pienin, monomeerinen sokerimolekyyli
syljen amylaasi: syljestä löytyvä ruoansulatusentsyymi, joka aloittaa suun hiilihydraattien pilkkoutumisen
soluhengitys: ATP: n tuotanto glukoosin hapettumisesta glykolyysin, Krebs-kierron ja oksidatiivisen fosforylaation kautta
glykolyysi: metabolisten reaktioiden sarja, joka hajottaa glukoosin pyruvaatiksi ja tuottaa ATP: tä
pyruvaatti: glykolyysin kolmihiilinen lopputuote ja lähtöaine, joka muuttuu asetyyli-CoA: ksi, joka tulee
Krebs-sykliin: kutsutaan myös sitruunahappokierroksi tai trikarboksyylihappokierroksi, muuntaa pyruvaatin CO2-ja suurenergisiksi fadh2 -, NADH-ja ATP-molekyyleiksi
sitruunahappokierroksi tai trikarboksyylihappokierroksi (TCA): kutsutaan myös Krebs-sykliksi tai trikarboksyylihappokierroksi; muuntaa pyruvaatin CO2-ja suurenergisiksi FADH2 -, NADH-ja ATP-molekyyleiksi
energiaa kuluttava vaihe, glykolyysin ensimmäinen vaihe, jossa tarvitaan kaksi ATP-molekyyliä aloita reaktio
glukoosi-6-fosfaatti: glykolyysin ensimmäisessä vaiheessa tuotettu fosforyloitu glukoosi
heksokinaasi: useimmissa kudoksissa esiintyvä soluentsyymi, joka muuttaa glukoosin glukoosi-6-fosfaatiksi soluunoton yhteydessä
Glukokinaasi: maksassa esiintyvä sellularentsyymi, joka muuttaa glukoosin glukoosi-6-fosfaatiksi soluunoton yhteydessä
energiaa tuottava vaihe: glykolyysin toinen vaihe, jonka aikana syntyy energiaa
terminaalinen elektroniseptori: ATP: n tuotantotie, jossa elektronit kulkevat hapetus-pelkistysreaktioiden sarjan läpi, joka muodostaa vettä ja tuottaa protonigradientin
Krebsin sykli: kutsutaan myös sitruunahappokierroksi tai trikarboksyylihappokierroksi, muuntaa pyruvaatin CO2: ksi ja suurienergisiksi FADH2 -, NADH-ja ATP-molekyyleiksi
elektroninsiirtoketju (jne.): ATP: n tuotantoreitti, jossa elektronit kulkevat hapetus-pelkistysreaktioiden sarjan läpi, joka muodostaa vettä ja tuottaa protonigradientin
oksidatiivinen fosforylaatio: prosessi, joka muuttaa suurienergisen NADH: n ja FADH2: n ATP: ksi
ATP: n syntaasiproteiini: huokoskompleksi, joka luo ATP: tä
p>
glukoneogeneesi: glukoosisynteesi pyruvaatista tai muista molekyyleistä