Introduction
fossiiliset luonnonvarat ovat energian ja orgaanisten yhdisteiden tärkein raaka-aine, ja niiden käytöstä aiheutuu ilmastonmuutokseen liittyviä kasvihuonekaasupäästöjä. Tuleva ilmastoromahdus edellyttää pikaista siirtymistä fossiilipohjaisesta biotalouteen, jossa polttoaineiden, kemikaalien ja materiaalien tuotantoon käytetään öljyn sijaan lignoselluloosamaista biomassaa. Puu on tärkeä lignoselluloosabiomassan lähde; se koostuu pääasiassa sekundaarisesti paksuuntuneista soluseinistä, joissa on runsaasti selluloosaa, hemiselluloosaa ja ligniiniä. Kaikki kolme polymeeriä voidaan hyödyntää biotaloudessa. Selluloosa on sellu-ja paperiteollisuuden lähde, ja sekä selluloosa-että hemiselluloosat voidaan depolymeroida monosakkarideikseen käymiseen esimerkiksi bioetanoliin, maitohappoon ja pesuaineisiin (Vanholme et al., 2013b). Koska ligniini vaikuttaa kielteisesti puunjalostuksen tehokkuuteen näihin käyttötarkoituksiin nähden, puut voidaan rakentaa siten, että ligniiniä kertyy vähemmän, jolloin se soveltuu paremmin paperin ja käymiskelpoisten sokereiden tuotantoon. Toisaalta ligniiniä pidetään yhä enemmän arvokkaana osana biotaloutta. Koska ligniini on maapallon suurin uusiutuva aromaattinen lähde, biojalostamon taloudellinen kannattavuus voi kasvaa merkittävästi, jos ligniiniä käytetään myös valoresurssina kemikaalien tuotannossa (Holladay ym., 2007; Tuck et al., 2012; Davis ym., 2013; Ragauskas et al., 2014; Li C. et al., 2015; Van den Bosch et al., 2015; Rinaldi ym., 2016; Upton and Kasko, 2016; Schutyser et al., 2018).
ligniinipolymeeri koostuu monolignoleista, joita tuotetaan fenyylipropanoidin ja monolignolin biosynteettisten reittien kautta sarjalla entsymaattisia reaktioita, jotka alkavat fenyylialaniinin deaminaatiosta (Kuva 1). Monolignolit syntetisoidaan sytoplasmassa ja translokoituvat apoplasmaan, jossa ne dehydrogenoituvat monolignoliradikaaleiksi lacaces-ja peroksidaasien vaikutuksesta (Berthet et al., 2011; Zhao et al., 2013). Nämä monolignoliradikaalit sitten pariutuvat keskenään kombinatorisesti, jolloin syntyy erilaisia kemiallisia sidoksia, kuten aryylieetterisidos (β-O-4), resinolisidos (β-β) ja fenyylikumaraanisidos (β-5) (Boerjan et al., 2003; Ralph et al., 2004; Vanholme ym., 2010). Yleisimpiä monolignoleja ovat hydroksikinnamyylialkoholit p-kumaryyli, koniferyyli ja sinapyylialkoholit, jotka tuottavat H -, G-ja S-yksiköt ligniinipolymeeriin lisättyään (Bonawitz and Chapple, 2010; Ralph et al., 2019; Vanholme ym., 2019). Ligniinin rakenneosien suhteellinen osuus vaihtelee taksonien, kehitysvaiheen, kudoksen ja solutyypin ja jopa soluseinän kerroksen välillä; havupuiden ligniini (gymnospermit) koostuu lähes kokonaan G-yksiköistä, joissa on pieni murto-osa H-yksiköistä, kun taas lehtipuiden ligniinillä (koppisiemeniset) on G-yksiköiden lisäksi s-yksiköitä ja H-yksiköiden jälkiä (Boerjan et al., 2003; Vanholme ym., 2010, 2019). Näiden perinteisten monolignolien lisäksi ligniinipolymeeriin voidaan liittää useita muita P-hydroksyloituja aromaattisia molekyylejä eri tasoille (Vanholme et al., 2019).
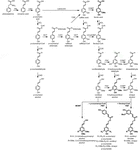
Figure 1. Lignin biosynthetic pathway. Alternative monomers and heterologously expressed enzymes are shown in bold. PAL, phenylalanine ammonia-lyase; C4H, cinnamate 4-hydroxylase; 4CL, 4-coumarate:CoA ligase; HCT, p-Hydroxycinnamoyl-CoA:quinate/shikimate-p-hydroxycinnamoyltransferase; C3’H, p-coumaroyl quinate/shikimate 3’-hydroxylase; CSE, caffeoyl shikimate esterase; CCoAOMT, caffeoyl-CoA O-methyltransferase; CCR, cinnamoyl-CoA reductase; F5H, ferulate 5-hydroxylase/CAld5H, coniferaldehyde 5-hydroxylase; COMT, caffeiinihappo-O-metyylitransferaasi; CAD, kinnamyylialkoholidehydrogenaasi; FMT, feruloyyli-CoA-monolignolitransferaasi; PMT, P-kumaroyyli-CoA-monolignolitransferaasi.
koska ligniini on merkittävä resalsitrianssitekijä puun delignifikaatioprosesseissa, ligniinin biosynteettisen reitin purkamiseen ja ligniinin biosynteesigeenien häiriöiden vaikutuksiin ligniinin määrään ja koostumukseen sekä puunjalostustehokkuuteen on panostettu runsaasti tutkimustoimintaa. Vaikka muutokset geeneissä, jotka vaihtelevat transkriptiotekijöistä oksidatiivisia entsyymejä koodaaviin tekijöihin, ovat johtaneet ligniinipitoisuuden, koostumuksen tai laskeuman muuttumiseen (Eriksson et al., 2000; Li Y. H. ym., 2003; Liang et al., 2008; Lu et al., 2013; Lin et al., 2016; Xu et al., 2017; Yang et al., 2017; Obudulu et al., 2018), tässä katsauksessa keskitytään ligniinin biosynteettisten geenien suunnittelun tuloksiin.
Ligniinireitin suunnittelu
taulukossa 1 esitetään yleiskatsaus poppelissa, männyssä, eukalyptuksessa ja koivussa esiintyviä alasääteltyjä tai mutatoituneita ligniinin biosynteettisiä geenejä koskevista eri tutkimuksista ja niiden vaikutuksista puunjalostustehokkuuteen määritettynä. Ligniinin biosynteettisen reitin minkä tahansa vaiheen aktiivisuuden vähentäminen PAL: stä CAD: hen voi johtaa ligniinipitoisuuden vähenemiseen (Taulukko 1). Ligniinin pelkistymisasteeseen vaikuttavat useat muuttujat, kuten kohdegeeni ja entsyymiaktiivisuuden alasääntelyn aste, joka puolestaan riippuu käytetyn äänenvaimennuskonstruktion tehokkuudesta, geeniperheen koosta ja redundanssista geeniperheen sisällä. Yleensä c4h: sta CCR: ään johtavien vaiheiden vähentäminen johtaa ligniinin määrän dramaattisempaan vähenemiseen (Hu et al., 1999; Meyermans et al., 2000; Zhong et al., 2000; Li L. et al., 2003; Jia et al., 2004; Lu ym., 2004; Leplé et al., 2007; Coleman ym., 2008a, b; Bjurhager et al., 2010; Mansfield ym., 2012; Ralph et al., 2012; Min et al., 2014; Van Acker ym., 2014; Zhou ym., 2015, 2018; Saleme et al., 2017; Xiang et al., 2017) kuin f5h: n, COMT: n ja CAD: n alasääntely (Van Doorsselaere et al., 1995; Baucher et al., 1996; Lapierre ym., 1999; Jouanin et al., 2000; Van Acker ym., 2014; Wang et al., 2018). Ligniinin pelkistyminen voi liittyä S/G: n lisääntymiseen, kuten C3 ’ H- (Coleman et al., 2008a; Ralph et al., 2012)ja CCoAOMT-downregulated trees (Meyermans et al., 2000), tai lasku S / G suhde kuten CSE – (Saleme et al., 2017), ja COMT-downregulated trees (Van Doorsselaere et al., 1995; Lapierre ym., 1999; Jouanin et al., 2000). Mielenkiintoista on, että low-lignin 4CL-downregulated poppelit havaittiin olevan kasvua S/G (Min et al., 2014; Xiang et al., 2017), s/G: n lasku (Voelker et al., 2010; Zhou ym., 2015) eli villityyppiin verrattavia suhdelukuja (Hu et al., 1999; Li L. et al., 2003). Tätä varianssia ei voida yhdistää promoottoriin tai alasäätelyssä käytettyyn menetelmään, mikä viittaa siihen, että äänenvaimennusasteen, kasvuolosuhteiden tai kehitystilan erot vaikuttavat tähän ominaisuuteen. Toisaalta voimakkaimpia vaikutuksia H / G / s ligniinikoostumukseen on havaittu C3 ’ h: n ja HCT: n alaskirjatuilla puilla, jotka tallettavat ligniiniä tuntuvasti H-yksikköpitoisuuden noustessa (Coleman et al., 2008a; Ralph et al., 2012; Vanholme ym., 2013a), kun taas puut, jotka yli-ilmentävät F5H: ta, tuottavat voimakkaasti s-yksiköissä rikastettua ligniiniä (Franke ym., 2000; Li L. et al., 2003; Stewart et al., 2009), ja puut, jotka ovat alasasetellaan COMT ovat dramaattisesti vähentäneet s yksikkö biosynteesi (Van Doorsselaere et al., 1995; Lapierre ym., 1999).

Taulukko 1. Katsaus metsäpuihin, joilla on ligniinin biosynteesigeenien modifioitu ilmentymä.
biomassan käsittelytehokkuutta voi muuttaa myös se, että luonnonvaraisten kasvien ligniinissä on enemmän molekyylejä, jotka yleensä ovat vähäisiä aineosia. Ferulihapon liittäminen CCR-puutoksiin johtaa asetaalisidosten muodostumiseen ligniinipolymeerissä, joka pilkkoutuu helposti happamissa biomassan esikäsittelyissä (Leplé et al., 2007; Ralph et al., 2008; Van Acker ym., 2014). Itse asiassa ligniinin ferulihappopitoisuudet korreloivat positiivisesti korkeamman sakarointitehokkuuden kanssa (Van Acker ym., 2014). 5-hydroksikoniferyylialkoholin ja 5-hydroksikoniferaldehydin liittäminen komt-puutosten (Van Doorsselaere et al., 1995; Lapierre ym., 1999; Jouanin et al., 2000; Morreel ym., 2004; Lu ym., 2010) synnyttää bentsodioksaanisidoksia, jotka mahdollisesti estävät kovalenttisia sidoksia ligniinin ja polysakkaridihydroksyyliryhmien välillä (Weng et al., 2010; Vanholme ym., 2012; Nishimura et al., 2018). Toisaalta COMT-puutos johtaa myös kondensoituneempaan ligniiniin, koska kondensoituneet β-β-Ja β-5-sidokset ovat suhteellisesti suurempia ja β-O-4-sidokset matalampia, kun S-yksikkötaajuus laskee. Poppeleista saadun puun kemiallinen massatuotanto, jota on voimakkaasti vähennetty COMT: tä varten, johti suurempaan sellutuottoon, jota tasapainottaa massan ligniinijäämä. Näissä puissa oli pienempi ligniini ja korkeampi selluloosapitoisuus (Jouanin et al., 2000). Toisaalta poppeleilla, joita säänneltiin COMT: n osalta vaatimattomasti, sellun tuotto laski huomattavasti, oletettavasti siksi, että ligniinipitoisuus oli pysynyt normaalina, kun taas ligniinissä oli enemmän tiivistyneitä sidoksia, jotka vaikuttivat negatiivisesti ligniinin uuttoon (Lapierre et al., 1999; Pilatus ym., 2002). Kinnamaldehydien liittäminen ligniinipolymeeriin CAD-puutoksissa johtaa lyhyempiin ligniinipolymeeriketjuihin, jolloin vapaiden fenolisten pääteryhmien osuus kasvaa, mikä lisää polymeerin liukoisuutta emäkseen. Kinnamaldehydien liittäminen ligniinipolymeeriin oletettavasti myös vähentää alifaattisen ketjun kovalenttista vuorovaikutusta hemiselluloosan kanssa, jolloin ligniini on jälleen liukoisempi. Lisäksi kanelialdehydin β-O-4: n pariutuessa toisen monomeerin kanssa syntyy pidennetyn konjugoituneen systeemin vuoksi sisältyvän kanelialdehydin aromaattinen eetterisidos herkistyy emäksiselle pilkkoutumiselle (Lapierre et al., 1989; Van Acker ym., 2017).
Ligniinipolymerointi on kombinatorinen radikaalikytkentäprosessi, jossa ligniinipolymeeriin voi luontaisesti liittyä monenlaisia fenoliyhdisteitä (Boerjan et al., 2003; Vanholme ym., 2019). Tutkijat ovat pyrkineet räätälöimään ligniinin määrää ja koostumusta biomassan prosessoinnin parantamiseksi heterologisten geenien ilmentymisen avulla. tavoitteena on biosynteesi ja erilaisten yhteensopivien fenoliyhdisteiden liittäminen vaihtoehtoisina monolignoleina ligniinipolymeeriin (Ralph, 2006; Vanholme et al., 2012; Mottiar ym., 2016; Mahon and Mansfield, 2018). Yksi esimerkki on käyttöön geenien koodaus entsyymejä, joita tarvitaan s yksikkö biosynteesi männyn; samanaikainen ilmentyminen F5H, COMT ja CAD onnistuneesti käyttöön s yksiköt Pinus radiata (Wagner et al., 2015; Edmunds et al., 2017). Monolignoli-4-O-metyylitransferaasia (MOMT4) koodaavan geenin kulkeutuminen poppeliin johtaa 4-o-metyloitujen havuperyyli-ja sinapyylialkoholien muodostumiseen, joita ei voida liittää kasvavaan ligniinipolymeeriin, koska niiltä puuttuu aromaattinen hydroksyyliryhmä. Tämä johtaa ligniinin polymeroitumisen pysähtymiseen ja johtaa puihin, joiden ligniinipitoisuus on pienempi ja sakarointitehokkuus suurempi (Bhuiya and Liu, 2010; Cai et al., 2016). Poppelit on myös suunniteltu sisältämään esterisidoksia ligniinipolymeerin selkärangassa. Coniferyyliferulaattiesterit kulkeutuivat polymeeriin Angelica sinensis (Wilkerson et al., 2014), mikä parantaa sakaroinnin tehokkuutta erilaisissa esikäsittelyolosuhteissa (Wilkerson et al., 2014; Kim et al., 2017; Bhalla et al., 2018), ja parannettu Kraftin sellutehokkuus verrattuna wild-tyyppiin (Zhou et al., 2017). Monolignoli-P-kumaraattiestereitä on myös kehitetty poppeleissa riisin p-KUMAROYYLI-CoA:monolignolitransferaasi (PMT) – geenin ilmentymisen kautta, jolloin resistenttien interunit-sidosten taajuus on suurempi ja G-ja s-terminaaliyksiköt vapailla fenoliryhmillä (Smith et al., 2015; Sibout et al., 2016). Arabidopsiksessa PMT: n heterologinen ilmentyminen johti ligniinin määrän vähenemiseen ja sakaroinnin tehostumiseen (Sibout et al., 2016), poppelin ligniinimäärässä ei havaittu laskua eikä sakaroinnin tehokkuutta määritetty (Smith ym., 2015).
vaikka useiden ligniinin määrää ja koostumusta koskevien muutosten osoitettiin parantavan biomassan käsittelyä, näihin muutoksiin liittyi usein biomassan saantoseuraamus (Leplé et al., 2007; Wadenbäck ym., 2008; Wagner ym., 2009; Voelker et al., 2010; Stout et al., 2014; Van Acker ym., 2014; Sykes et al., 2015; Zhou et al., 2018). Äskettäin levinnyt metastudia häiritsi 21 ligniinin biosynteesigeenin P. trichocarpa, ja yhdisti kattavasti transkriptomisten, proteomisten, fluksomisten ja fenomisten 221 rivin tulosten tulokset. Kirjoittajat päättelivät, että puiden kasvuun ei liity ligniinin määrää, alayksiköiden koostumusta tai erityisiä yhteyksiä (Wang et al., 2018), mutta pikemminkin korreloi romahtaneiden xylem-alusten läsnäolon kanssa (Coleman et al., 2008a, b; Wagner et al., 2009; Voelker et al., 2010; Vargas et al., 2016; De Meester et al., 2018), soluseinän eheysreitin aktivointi (Bonawitz et al., 2014)ja / tai kemiallisten inhibiittorien kertyminen (Gallego-Giraldo et al., 2011; Muro-Villanueva ym., 2019).
vaikka ligniinipitoisuutta on pyritty merkittävästi vähentämään biosynteettisten ligniinigeenien säätelyn vähentämisellä, ligniinireitin säätelyä ja ligniinin ylituotantoa koskevat tutkimukset ovat olleet vähissä. Itse asiassa raportit F5H: n yliekspressiosta osoittavat, että ligniinipitoisuus on ennallaan tai jopa vähentynyt (Huntley et al., 2003; Li L. et al., 2003; Stewart et al., 2009; Mansfield ym., 2012; Edmunds et al., 2017). CAD: n ja COMT: n yliekspressio on johtanut pikemminkin geenien hiljentymiseen kuin säätelyyn, tai mitään vaikutusta ekspressiotasoihin ei havaittu (Baucher et al., 1996; Lapierre ym., 1999; Jouanin et al., 2000; Leplé et al., 2007; Van Acker ym., 2014). R2r3-MYB-transkriptiotekijöiden PtoMYB92, PtoMYB216 ja PtoMYB74 yliekspressio johti kaikki ylimääräisiin ksyleemikerroksiin, paksumpiin ksyleemisoluseiniin sekä ektooppiseen ligniinikerrostumaan, ja kasveihin kertyi 13-50% enemmän ligniiniä (Tian Q. et al., 2013; Li C. F. et al., 2015; Li et al., 2018). MYB: n yliekspressiolinjat säätelivät konstitutiivisesti ligniinin biosynteesireitin geenejä, ja vaikka myb92: n ja MYB74: n yliekspressiolla oli biomassarangaistus, MYB216: n yliekspressio johti kasveihin, joissa oli jopa 50% enemmän ligniiniä eikä lainkaan kehittyvää fenotyyppiä. Koska ligniiniä pidetään yhä enemmän tärkeänä resurssina kemikaalien kestävälle tuotannolle (Cao et al., 2018) ligniiniä liikaa tuottavien laitosten suunnittelua tulisi tutkia tarkemmin.
kenttäkokeet
edellä käsitellyt esimerkit osoittavat selvästi, että ligniinin valmistaminen fenyylipropanoidireittigeenien alas – tai yläsääntelyn avulla – tai heterologisten geenien ilmentymisen avulla – voi lisätä lignoselluloosabiomassan prosessointitehokkuutta. Käytännön ja sääntelyn vuoksi useimmat tutkimukset raportoivat kasvihuoneessa kasvatettujen puiden analyysistä saaduista tiedoista. Kasvihuoneessa kasvatetuilla puilla tehdyissä kokeissa ei kuitenkaan yleensä oteta huomioon kehitysprosesseja, kuten kasvun pysähtymistä ja lepotilaa. Kasvihuonekokeet eivät myöskään anna riittävää tietoa muokatun kasvin vuorovaikutuksesta ympäristötekijöiden, kuten maalajin, tuulen ja taudinaiheuttajien kanssa. Näiden vuorovaikutusten ymmärtäminen on tärkeä askel tutkimustulosten kääntämisessä kaupallisiin sovelluksiin. Tutkimukset, joille myönnettiin lupa kenttäkokeiden laatimiseen, tuovat esiin merkittäviä fenotyyppieroja kasvihuonepuiden ja pellolla kasvatettujen puiden välillä. Taulukossa 1 on yhteenveto 4CL -, CCoAOMT -, CCR -, COMT-ja CAD-downregulated-puilla tehdyistä kenttäkokeista.
useiden lignoselluloosapuiden potentiaalin käyttökokeiden substraattina vahvistivat useat ligniinitekniikalla kasvatetut pellolla kasvatetut puut, jotka osoittivat puunjalostuksen parantuneen. Poppelit, joita on vähennetty Ccoaomtille, kasvatettu 5 vuotta kenttäkokeessa Pekingissä (Kiina), osoittivat lisääntyneen glukoosin ja ksyloosin vapautumisen sakaroinnin yhteydessä (Wang et al., 2012). Poppelit, joita säädeltiin CCR: ään ja joita kasvatettiin kenttäkokeessa Ranskassa, osoittautuivat soveltuvammiksi kemialliselle kraftmassaukselle (Leplé et al., 2007). Kaksi muuta KENTTÄKOETTA, jotka tehtiin Ranskassa ja Belgiassa CCR-downregulated poppeleilla, paransivat etanolin tuotantoa 160%: lla samanaikaisissa sakarointi-ja fermentointimäärityksissä (SSF); laitosten biomassa väheni kuitenkin jopa 50% (Van Acker et al., 2014). Myös kenttäkokeet CAD-downregulated poppelilla osoittivat lupaavia tuloksia. Näissä puissa oli hieman vähemmän ligniiniä kuin villityypeissä ja ne osoittautuivat halukkaammiksi Kraftin delignifikaatioon (Lapierre et al., 1999). Ranskassa ja Yhdistyneessä kuningaskunnassa tehdyissä suuremmissa kenttäkokeissa samoilla linjoilla ligniinin määrä väheni lievästi ja kaupallisesti merkityksellisenä pidetty Kraftin sellun määrä parani, koska kasvit tarvitsivat 6% vähemmän emästä, jotta saataisiin aikaan samanlainen delignifikaatio kuin luonnonvaraisissa puissa (Pilatus et al., 2002).
ristiriitaiset raportit sekä biomassatuotoksesta että jatkojalostuksen tehokkuudesta viittaavat kuitenkin siihen, että ympäristötekijät vaikuttavat suuresti näihin parametreihin. Kiinassa tehdyssä kenttätutkimuksessa, jossa käytettiin 4CL: n alasäädeltyjä poppeleita, havaittiin, että vaikka ligniinipitoisuus pieneni 28% villityyppiin verrattuna, puiden korkeus kasvoi noin 8% (Tian X. M. et al., 2013), yhdenmukaisesti kasvihuone-tutkimusten (Hu et al., 1999), muissa kenttäkokeissa havaittiin, että 4CL-downregulated poppelit olivat vähentäneet biomassaa ja olivat joskus jopa kääpiöityjä (Voelker et al., 2010; Stout et al., 2014; Marchin ym., 2017). Raporteissa on eroja myös näistä 4CL-alasäädellyistä pellolla kasvatetuista poppeleista saadun puun jatkojalostustehokkuuden osalta. Vaikka jopa 100% kasvu sokerin talteenotto todettiin 4cl1-alasäädelty puita (35S-ajettu antisense 4CL rakentaa)kasvanut vuoristoalueella Yhdysvalloissa (Xiang et al., 2017), Oregonissa (Yhdysvalloissa) tehdyistä kenttätutkimuksista saadut tiedot osoittivat, että Pt4CL1 promoottori-driven antisense silenced 4CL1 poppelit eivät parantaneet sakaroinnin tehokkuutta verrattuna villityyppiin (Voelker et al., 2010). Samoin pitkän aikavälin tutkimus Wenling (Kiina), totesi, että 4CL-downregulated poppelit eivät osoittaneet merkittävää parannusta sokerin saanto verrattuna villi tyyppi (Wang et al., 2012). Molemmissa viimeksi mainituissa tapauksissa puut vähenivät ligniinin määrää lievästi, mikä ei johtanut korkeampaan jalostustehokkuuteen, mikä saattaa johtua entsymaattiseen aktiivisuuteen mahdollisesti vaikuttavien uuteaineiden korkeammasta pitoisuudesta (Voelker ym., 2010).
kenttätutkimuksissa on osoitettu, että ympäristötekijät voivat vaikuttaa lignifikaatioon ja palauttaa luonteenpiirteet villin tyyppisille tasoille verrattuna tasoihin, jotka saavutettiin, kun samoja kasveja kasvatettiin kasvihuoneessa. Vaikka 4CL-alasäädetyt puut olivat vähentäneet ligniinipitoisuutta kasvihuoneolosuhteissa kasvatettaessa, samojen 4CL-antisensiteettipoppeleiden, mutta pellolla kasvatettujen, analyysi on usein osoittanut, että ligniinipitoisuus on kasvanut kasvihuoneessa kasvatettuihin puihin verrattuna ja joskus jopa palautettu villin tyypin tasolle (Stout et al., 2014; Xiang et al., 2017). Samoin LIGNIINIPITOISUUDET laskivat paljon vähemmän CCR-puutteellisissa poppeleissa, kun niitä kasvatettiin pellolla verrattuna kasvihuoneessa kasvatettuihin (Van Acker ym., 2014). Ainakin CCR-puutteellisten poppelien osalta on mahdollista, että peltokasvustojen korkeampi lignöitymisaste johtuu siitä, että puunäytteet on otettu talven aikana. Kun puun kasvu lakkaa syksyllä, puilla on vielä aikaa lignifioida soluseinänsä täysin puun lepotilaan mennessä, verrattuna kasvihuoneessa kasvaviin puihin, jotka kehittävät jatkuvasti uusia ksyleemejä. Ligniinikoostumuksen on myös osoitettu eroavan kasvihuone-ja peltokasveilla kasvavien matalien ligniinipuiden välillä. 4CL-downregulated poppelit kasvanut alalla Pohjois-Carolinassa oli ligniini pienempi S / G kuin silloin, kun samat linjat kasvatettiin kasvihuoneessa (Stout et al., 2014).
yhdessä nämä tulokset osoittavat, että kasvihuoneessa kasvatetuista puista saatuja tietoja ei voida helposti ekstrapoloida pellolla kasvatettuihin puihin, mikä tukee kenttäkokeiden tarvetta eri paikoissa. Jotkin linjat esittivät saantorangaistuksen, joka teki niistä vähemmän mielenkiintoisia sovelluksille, korostaen tarvetta ymmärtää paremmin saantorangaistuksen molekyylipohjaa ja kehittää strategioita tämän ongelman ratkaisemiseksi.
ligniinillä on osoitettu olevan tärkeä rooli taudinaiheuttajien resistenssissä (Miedes et al., 2014; Zhao and Dixon, 2014), ja sillä on keskeinen rooli siinä, että voimala voi kuljettaa vettä. Tämä viittaa siihen, että ligniinin muunnoksilla voi olla vaikutusta kasvien stressinsietokykyyn. Vaikka lisätutkimuksia tarvitaan, jotta tähän mahdollisuuteen voidaan täysin puuttua, 4CL: n, COMT: n ja CAD: n säätely poppelissa ei merkittävästi muuttanut lehtiä ruokkivien kasvinsyöjien ruokintakykyä (Tiimonen et al., 2005; Brodeur-Campbell ym., 2006; Hjalten et al., 2013). COMT: n ja CAD: n säätelyn vaikutusta kasvien ja hyönteisten vuorovaikutukseen poppelissa on arvioitu myös pellolla kasvatetuissa puissa, ja osoitettiin, että ligniinimuunnelluilla puilla oli normaali vierailevien ja ruokkivien hyönteisten esiintyvyys sekä normaali vaste mikrobipatogeeneille (Pilatus et al., 2002; Halpin ym., 2007). Tulokset viittaavat siihen, että muuntunutta ligniiniä sisältävät puut eivät välttämättä kärsi tuholaisista ja taudeista luonnonvaraisia kasveja enempää. Kuitenkin profilointi endosfäärin bakteerien mikrobiomin korjatun puun pellolla kasvatetuista, CCR-alasäädellyistä poppeleista osoitti muutoksia bakteeri yhteisössä, oletettavasti siksi, että tiettyjen fenolisten metaboliittien muuttunut runsaus xylem (Beckers et al., 2017).
ottaen huomioon ligniinin roolin ksyleemin toiminnassa ja rakenteessa on arvioitu muutamien matalien ligniinimuunneltujen poppeleiden vesisuhteita. 4CL-downregulated poppelit havaittiin olevan alentunut hydraulinen johtavuus, mahdollisesti häiritsee kasvien kasvua (Marchin et al., 2017). Hydraulinen stressi kokeet poppelit downregulated CCR, COMT tai CAD osoitti, että nämä kasvit oli pienempi vastustuskyky kavitaatio, säilyttäen normaalin xylem hydraulinen johtavuus ja veden kuljetus (Awad et al., 2012). Tulokset viittaavat siihen, että vähäligniinimutanttien kasvuun saattaa vaikuttaa veden saatavuus. Kuten minkä tahansa klassisesta jalostuksesta saadun uuden hybridin kohdalla, tarvitaan kenttätestejä, jotta voidaan arvioida ligniinillä muokattujen puiden kenttätehokkuutta ja stressinsietokykyä.
näkymät Ligniinitekniikan kehittymiselle Metsäpuissa
ympäristöolosuhteet näyttävät vaikuttavan suuresti ligniinituotantolaitosten suorituskykyyn. On kuitenkin epäselvää, johtuvatko kasvihuoneessa kasvatettujen ja pellolla kasvatettujen puiden tai eri peltoalueilla kasvatettujen puiden väliset erot geenien suppression eritasoista vai muokatun ominaisuuden vuorovaikutuksesta ympäristön kanssa (GxE). Itse asiassa epävakaa downregulation on puute geenien hiljentämiseen tekniikoita, jotka perustuvat RNAi. Tämä on todistettavissa havainnoimalla punaisen ksyleemin fenotyypin vaihtelua, joka havaitaan, kun tietyt ligniinin biosynteesigeenit, kuten CAD, COMT tai CCR, ovat downregulated. Punainen xylem väri ei usein ole yhtenäinen koko xylem, vaan esiintyy laikkuja, jotka heijastavat vaihtelevia tasoja geenin hiljentymistä (Leplé et al., 2007; Voelker et al., 2010; Van Acker ym., 2014; kuva 2). Lisäksi geenien vaientamismenetelmien käyttö voi mahdollisesti johtaa siihen, että lähisukulaisten geeniperimän jäsenten samanaikainen vaientaminen – ehkä eriasteisesti – sumentaa yksittäisten geenien tulkintoja ja naamioi niiden vaikutukset.

kuva 2. RNAi: n hajanainen gene downregulation. Kirjava punainen ksyleemi-fenotyyppi, joka on havaittu Belgiassa kenttäkokeessa kasvatettujen CCR-puutteellisten poppelien rungoissa (oikealla). Punainen ksyleemi osoittaa CCR: n alasääntelyn alueita. Luonnontyyppisten puiden puu on vaaleaa (vas.).
nämä ongelmat voidaan nyt helposti ratkaista käyttämällä CRISPR-pohjaisia geenimuokkaustekniikoita, jotka mahdollistavat tiettyjen kohdegeenien stabiilit funktionaalisen mutaation (knock-out), jolloin yksittäisten geenien toiminta voidaan dissektiolla perheiden sisällä. Esimerkiksi yksittäisten 4CL-geeniperheen jäsenten kohdentaminen poppeliin osoitti, että 4cl1 liittyy lignifikaatioon, kun taas 4CL2 osallistuu proantosyanidiinin tuotantoon (Zhou et al., 2015). Knock-out-alleelien lisäksi CRISPR-pohjainen geenieditointi mahdollistaa myös uusien alleelien luomisen, jotka heikentävät entsyymiaktiivisuutta osittain. Tämä antaa mahdollisuuden hienosäätää entsyymiaktiivisuuden määrää ja ohittaa saantoseuraamus, joka usein havaitaan ligniinin määrän laskiessa kynnysarvon alapuolelle. Toinen lupaava väylä metsäpuiden ligniinitekniikalle, joka on mahdollista CRISPR-pohjaisen genomitekniikan avulla, on useiden geenien (alleelien pinoaminen) samanaikainen muokkaus biomassan käsittelyn tehokkuuden optimoimiseksi, kuten Arabidopsis-julkaisussa, jossa transaldolaasin (tra) ja comt-mutaatioiden pinoaminen, c4h-ja comt-mutaatiot tai 4cl-ja comt-mutaatiot johtivat additiivisiin ja synergistisiin parannuksiin sakaroinnin tehokkuudessa (de Vries et al., 2018). Itse asiassa a systems approach in P. trichocarpa ennustaa, että pal: n ja Ccoaomtin eli PAL: n, C3 ’ h: n ja CCOAOMTIN samanaikainen alasääntely parantaa merkittävästi puun ominaisuuksia ja sokerin vapautumista (Wang et al., 2018).
CRISPR-pohjaisen genomieditoinnin käyttö puunparannuksessa sellu-ja paperiteollisuudessa sekä biojalostamoteollisuudessa sekä ligniinin hydrogenolyyttisestä hajoamisesta syntyvien alustaromaattien tuotannossa on arvokkainta, kun tämä tekniikka yhdistetään strategisesti muihin jalostustekniikoihin (kuva 3). Ligniinin määrässä ja S/G-koostumuksessa on jo suuria vaihteluja metsäpuiden luonnollisissa populaatioissa (Studer et al., 2011). Koska molemmat ominaisuudet vaikuttavat glukoosin vapautumiseen sakaroinnin yhteydessä (Yoo et al., 2018), tämän geneettisen monimuotoisuuden hyödyntäminen tavanomaisella jalostuksella, apuna Genome Wide Association Studies (Gwas) (Porth et al., 2013; Fahrenkrog et al., 2017; Liu et al., 2018), harvinaisten viallisten alleelien (BRDA) jalostus (Vanholme et al., 2013a) tai genomivalinta (Yin et al., 2010; Muchero et al., 2015; Pawar et al., 2018; Xie et al., 2018), on arvokas strategia saada linjoja, jotka ovat parantaneet puunjalostuksen tehokkuutta. Kun eliittipuut saadaan näillä jalostusmenetelmillä, tiettyjen geenien geenitekniikka ja CRISPR-pohjainen geenieditointi on erittäin lupaava väylä parantaa edelleen näitä eliittigenotyyppejä rikkomatta niiden geneettistä rakennetta ja käymättä läpi pitkiä jalostusjaksoja. Koska ilmasto romahtaa pian, meillä ei ole enää aikaa hukattavana näiden uusien jalostustekniikoiden omaksumisessa kilpajuoksussamme biopohjaiseen talouteen.
kuva 3. Metsäpuiden geneettinen parantaminen jalostusvälineiden yhdistelmällä. Jotta metsäpuiden geneettistä parantumista sellu-ja biojalostamosovelluksiin voidaan nopeuttaa, klassiset ja uudet jalostusvälineet on yhdistettävä fiksusti. Klassinen jalostus tarkoittaa puiden fenotyyppistä valintaa kontrolloituihin risteytyksiin, minkä jälkeen seuraa fenotyyppinen valinta. Monien metsäpuiden genomisekvenssitietojen myötä on kehitetty uusia strategioita, kuten Genomivalinta, genomin laaja Assosiaatiotutkimus (Gwas) ja harvinaisten viallisten alleelien jalostus (BRDA), joilla nopeutetaan hyödyllisiin ominaisuuksiin liittyvien DNA-polymorfismien talteenottoa ja rikastamista. CRISPR-pohjainen genomieditointi mahdollistaa genomin muokkaamisen tavalla, joka jäljittelee luonnollisia polymorfismeja. Geenimuunteluun kuuluu vieraan DNA: n stabiili integrointi puuhun tuottamaan (an) entsyymiä (- entsyymejä) tai downregulatoimaan (a) geeniä (- geenejä). Klassisen ja uuden jalostustekniikan yhdistäminen on välttämätöntä, jotta yhteiskunnalle saadaan riittävästi korkealaatuista puuta.
Tekijäosuudet
kaikki luetellut tekijät ovat antaneet merkittävän, suoran ja älyllisen panoksen teokseen ja hyväksyneet sen julkaistavaksi.
Rahoitus
hyväksymme osittaisen rahoituksen IWT-SBO-hankkeelta bioleum (apuraha nro 130039) ja SBO-FISHILTÄ ARBOREF-hankkeen kautta. AC on saanut rahoitusta FWO: lta ja Euroopan unionin Horisontti 2020-tutkimus-ja innovaatio-ohjelmalta Marie Skłodowska-Curie-avustussopimuksen nro 665501 mukaisesti. LdV: n rahoitti Flanderin Institute for the promotion of Innovation through Science and Technology (IWT-Vlaanderen) for a predoctoral fellowship.
Eturistiriitalausunto
kirjoittajat toteavat, että tutkimus tehtiin ilman kaupallisia tai taloudellisia suhteita, joita voitaisiin pitää mahdollisena eturistiriitana.
kiitokset
Kiitämme Annick Bleytä tämän käsikirjoituksen valmistelusta lähetettäväksi.
Boerjan, W., Ralph, J., and baucher, M. (2003). Ligniinin biosynteesi. Annu. Rev. Plant Biol. 54, 519–546.
PubMed Abstract/Google Scholar
Lapierre, C., Pollet, B., Petit-Conil, M., Toval, G., Romero, J., Pilatus, G., et al. (1999). Siirtogeenisissä poppeleissa olevien ligniinien rakenteellisilla muutoksilla, joilla on alentunut kinnamyylialkoholidehydrogenaasi tai kofeiinihapon O-metyylitransferaasiaktiivisuus, on päinvastainen vaikutus teollisen Kraftin sellun tehokkuuteen. Kasvifysioli. 119, 153–163.
PubMed Abstract/Google Scholar
Ralph, J. (2006). ”Mikä tekee hyvän monolignolin korvikkeen?, ”teoksessa The Science and Lore of the Plant Cell Wall – Biosynthesis, Structure and Function, toim. T. Hayashi (Boca Raton, FL: BrownWalker Press).
Google Scholar