Introduction
Une interaction réussie avec notre environnement sensoriel nécessite un équilibre complexe de deux mécanismes de sélection attentionnelle – celui de haut en bas et de bas en haut. En vous dirigeant vers l’allée des produits de votre supermarché local dans le but de ramasser peu d’ingrédients nécessaires à la salade de mangue, engagez le déploiement d’un système d’attention volontaire orienté vers l’objectif ou de haut en bas, de sorte que vous recherchez activement tous les ingrédients requis parmi la multitude de choix de produits. Cependant, si vous entendez une sonnerie d’un téléphone portable, elle captera probablement votre attention et interrompra votre recherche. Une telle interruption se produit d’une manière ascendante, ou impulsée par un stimulus, selon laquelle une simple saillance du stimulus, le fait que l’anneau est différent des autres sons de votre environnement, le juge digne de sélection. Le scénario décrit souligne l’importance de la sélection ciblée et impulsée par le stimulus pour le comportement, et indique un équilibre fin qui doit exister entre les deux systèmes attentionnels pour empêcher la « vision en tunnel” d’une part et l’incapacité totale de se concentrer sur l’autre.
Sélection de Haut en bas et de bas en haut: Comportement
Plusieurs décennies de recherche comportementale ont été consacrées à démontrer que la distribution de l’attention peut être contrôlée par les intentions de l’observateur ainsi que par la saillance du stimulus physique. Une grande partie des preuves comportementales de l’allocation attentionnelle descendante et ascendante ont été largement examinées ailleurs (Johnston et Dark, 1986; Egeth et Yantis, 1997). Pour résumer, les études démontrant les effets du contrôle attentionnel de haut en bas montrent que l’attention peut être allouée avec succès aux emplacements spatiaux, aux caractéristiques, aux objets, etc., à la suite de la présence de signaux exogènes ou endogènes (Eriksen et Hoffman, 1972; Posner, 1980; Posner et al.(Shaw, 1978; Moore et Egeth, 1998; Geng et Behrmann, 2002, 2005; Shomstein et Yantis, 2004a; Drummond et Shomstein, 2010). Les preuves à l’appui de l’allocation attentionnelle ascendante reposent sur divers paradigmes de capture attentionnelle, dans lesquels les participants sont engagés dans une recherche descendante et leur attention est détournée vers les stimuli non pertinents de la tâche, démontrant que l’attention est capturée par des singletons caractéristiques (unique item; Yantis et Jonides, 1990; Theeuwes, 1991; Folk et al., 2002) et des bouleversements brusques (Yantis et Jonides, 1984; Theeuwes, 1991; Koshino et al., 1992; Juola et coll., 1995).
Alors que la plupart des premières études se concentraient sur la démonstration de preuves de sélection attentionnelle descendante et ascendante, la plupart des études récentes se sont concentrées sur l’examen de l’interaction des deux systèmes de sélection attentionnelle. Cette ligne de recherche est alimentée par des observations selon lesquelles, pour sélectionner efficacement des informations pertinentes pour la tâche (par exemple, les ingrédients de la salade), il faut inhiber activement les informations non pertinentes pour la tâche qui détourneraient autrement l’attention de la tâche à accomplir. Le revers de cette logique est que moins on se concentre sur les informations liées aux tâches, plus la capture s’ensuivra. Il a été démontré expérimentalement que l’état attentionnel de l’observateur prédit quel type d’information, et dans quelle mesure, captera finalement l’attention (Folk et al., 1992, 2002; Bacon et Egeth, 1994; Gibson et Kelsey, 1998). Par exemple, Folk et al. (2002) ont montré que lors de la recherche d’une lettre rouge, un observateur sera plus facilement capturé par un stimulus non pertinent en périphérie si ce stimulus est rouge ou correspond d’une manière ou d’une autre au modèle cible. Étant donné que les paramètres de contrôle de haut en bas de l’observateur sont définis pour rechercher une caractéristique rouge, tout stimulus rouge est susceptible de capter l’attention et d’interférer potentiellement avec le contrôle de haut en bas. Ainsi, avec une tâche de capture, les stratégies de recherche attentionnelle peuvent être distinguées les unes des autres en faisant varier les niveaux de similarité entre les propriétés de stimulus de la cible et les distracteurs. Plus la cible est similaire au distracteur, plus il est difficile pour l’observateur d’éviter la capture.
Le rôle du Lobe pariétal dans la Sélection de Haut en Bas et de Bas en Haut: Neuroimagerie
Diverses techniques de neuroimagerie ont fourni des preuves solides de l’implication du cortex pariétal dans l’orientation descendante et ascendante, les preuves ayant été largement examinées ailleurs (Corbetta et Shulman, 2002, 2011; Behrmann et al., 2004). Il a été démontré que les zones les plus souvent activées suivant des indices de haut en bas pour assister à des emplacements, des caractéristiques ou des objets particuliers sont situées le long des parties dorsales du cortex pariétal. Ces zones comprennent le lobule pariétal inférieur (IPL), les régions dorsomédiales appelées lobule pariétal supérieur (SPL), ainsi que des régions plus médiales le long du gyrus précuneus (Yantis et al., 2002; Giesbrecht et coll., 2003; Liu et coll., 2003; Yantis et Serences, 2003; Figure 1). Plusieurs tâches descendantes ont été montrées pour engager avec succès des régions dorsales du cortex pariétal, à savoir celles impliquant l’espace (Kastner et al., 1999; Corbetta et coll., 2000; Hopfinger et coll., 2000; Shomstein et Behrmann, 2006; Chiu et Yantis, 2009; Greenberg et al., 2010) ainsi que des changements d’attention non spatiaux (Giesbrecht et al., 2003; Yantis et Serences, 2003; Shomstein et Yantis, 2004b, 2006; Tamber-Rosenau et al., 2011).
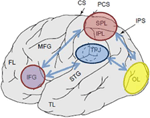
Figure 1. Représentation schématique de repères anatomiques pertinents projetés sur la surface latérale du cerveau humain. Le lobule pariétal supérieur (SPL) et le lobule pariétal inférieur (IPL) sont des régions situées dans la partie dorsale du cortex pariétal subissant une orientation attentionnelle de haut en bas. La jonction temporo-pariétale (TPJ) est une région du cortex pariétal ventral subissant une orientation attentionnelle ascendante. On pense que les deux, SPL et TPJ, suscitent des signaux de contrôle responsables des modulations attentionnelles ultérieures observées sur les régions sensorielles, dans ce cas la modulation (marquée par des flèches bleu foncé) de l’activité évoquée visuellement dans le lobe occipital (OL). De plus, on pense que les zones le long du gyrus frontal inférieur (IFG) et de la jonction frontale inférieure (IFJ) servent de zones de convergence pour le contrôle attentionnel piloté par le stimulus et de haut en bas (marquées par des flèches bidirectionnelles bleu clair).
Dans une tâche typique visant à engager l’allocation attentionnelle descendante, les individus reçoivent deux flux de présentation visuelle en série rapide (RSVP) positionnés en périphérie et sont initialement chargés de surveiller un flux pour un signal (par exemple, un chiffre parmi le flux de lettres). L’identité du signal indique si le sujet doit maintenir son attention sur le flux actuel ou déplacer son attention sur l’autre flux (Yantis et al., 2002; Yantis et Serences, 2003). Deux résultats majeurs sont observés dans de tels paradigmes. La première concerne une activation accrue dans les régions sensorielles représentant l’emplacement assisté en ce moment (par exemple, une activité accrue dans les régions visuelles primaires de gauche lorsque le flux RSVP de droite est suivi). Cette découverte fournit des preuves solides que les participants se rendent à un endroit précis et que l’attention module la force de la réponse sensorielle (voir Figure 1; Moran et Desimone, 1985; O’Craven et al., 1997). La deuxième découverte a trait à l’observation que les régions dorsales du lobe pariétal sont activées sélectivement par des changements d’attention de haut en bas. On observe que le cours temporel d’activité SPL/IPL est de nature transitoire suggérant que cette zone du cortex pariétal est la source d’un signal de commande attentionnel bref pour déplacer les états attentifs de manière descendante (Yantis et al., 2002).
Plusieurs études de l’IRMf ont documenté que la capture attentionnelle ascendante, médiée par la saillance et/ou la pertinence du stimulus, est maintenue par la jonction temporo-pariétale (TPJ; Figure 1). Par exemple, lorsque les sujets assistent et surveillent un changement d’un stimulus visuel ou auditif, présenté simultanément, l’activation des régions TPJ du lobe pariétal est améliorée. En plus de la sensibilité apparente aux stimuli pertinents, la TPJ est également activée en réponse à des événements potentiellement nouveaux (inattendus ou peu fréquents) lorsqu’un organisme est engagé dans un contexte comportemental neutre ou lorsqu’il est engagé dans une tâche (Marois et al., 2000; Downar et coll., 2002; Serences et coll., 2005; Corbetta et coll., 2008; Asplund et coll., 2010; Diquattro et Geng, 2011; Geng et Mangun, 2011). Cette activation se produit indépendamment de la modalité (auditive, tactile et visuelle) dans laquelle l’entrée est délivrée, reflétant la nature multisensorielle de la TPJ (mais voir Downar et al., 2001).
Dans une tâche typique examinant le mécanisme neuronal de la capture attentionnelle ascendante, les participants reçoivent un flux RSVP d’éléments au centre de l’écran et sont invités à identifier une cible prédéfinie (par exemple, identifier une lettre rouge présentée dans un flux RSVP de non-cibles blanches). Une certaine proportion des essais contient un distracteur saillant non pertinent à la tâche présenté à divers intervalles de temps avant le début de la cible, tandis que d’autres essais ne contiennent que le distracteur saillant (c’est-à-dire sans la cible). Des essais « Cible-distracteur » sont utilisés pour évaluer l’étendue de la capture, montrant que le distracteur non pertinent pour la tâche est en fait saillant, ce qui entraîne une diminution de la précision de la cible. Les essais de « distracteur isolé » sont utilisés pour des analyses plus poussées, car de tels essais permettent d’examiner l’activité provoquée par le distracteur saillant sans contamination par les processus liés à la cible. Plusieurs constats importants ressortent de ces paradigmes. Premièrement, lorsque les distracteurs sont séparés spatialement de l’emplacement cible, les distracteurs de capture s’accompagnent d’une activité corticale accrue dans les régions correspondantes du cortex sensoriel (par exemple, cortex visuel organisé rétinotopiquement; voir Figure 1). De tels résultats fournissent des preuves solides que lors de la capture, l’attention spatiale est en fait captée sur l’emplacement spatial occupé par le distracteur (Serences et al., 2005). Deuxièmement, les régions ventrales du cortex pariétal, principalement au sein de la TPJ, sont sélectivement activées par des changements d’attention involontaires de bas en haut. Tout comme l’activité au sein du SPL pour l’orientation descendante, le cours temporel d’activité observé sur TPJ est de nature transitoire suggérant que cette région est la source d’un bref signal de commande attentionnel pour déplacer l’attention de bas en haut.
Il convient de noter que bien que cette revue se concentre sur les fonctions cognitives du cortex pariétal postérieur, d’autres régions, notamment celles du cortex frontal, sont également recrutées pour une allocation attentionnelle de haut en bas et de bas en haut. Ces régions comprennent le cortex frontal ventral (VFC), les champs oculaires frontaux (FEF), la jonction frontale inférieure (IFJ) et le gyrus frontal inférieur (IFG; Corbetta et Shulman, 2002, 2011; Serences et al., 2005; Asplund et coll., 2010; Diquattro et Geng, 2011).
Le rôle du lobe pariétal dans la sélection de Haut en Bas et de Bas en haut: Neuropsychologie
Historiquement, les chercheurs se sont appuyés de manière critique sur des études neuropsychologiques de patients atteints de négligence hémispatiale (un trouble de l’allocation spatiale de l’attention à l’hémi-espace gauche) pour mieux comprendre les fonctions cognitives associées au lobe pariétal. Dans la littérature neuropsychologique classique, le cortex pariétal, dans son ensemble, était généralement considéré comme le site de lésion primaire de la négligence hémispatiale. Ce point de vue, élaboré en détail par les premiers chercheurs (Critchley, 1953; McFire et Zangwill, 1960; Piercy, 1964) ont clairement reconnu l’association entre la lésion pariétale et la négligence qui en a résulté. Cette perspective a été largement maintenue dans les années 1980 lorsque Posner et ses collègues (1984) ont utilisé le paradigme de repérage visuospatial secret pour montrer que les dommages au lobe pariétal produisent un déficit dans l’opération de « désengagement” (rétracter l’attention d’un endroit et la déplacer vers un autre) lorsque la cible est controlatérale à la lésion. Cependant, malgré cette avancée majeure dans la compréhension de la base neuronale de l’attention et en particulier du rôle de « désengagement” du cortex pariétal, leurs résultats supposent un seul site cortical (cortex pariétal) et une seule capacité fonctionnelle (« désengagement”). Contrairement à cette approche plus monolithique du cerveau (cortex pariétal) et du comportement (désengagement attentionnel), des travaux récents sur le comportement et la neuroimagerie (examinés ci-dessus et ailleurs) suggèrent que la région corticale et le comportement attentionnel associé peuvent être subdivisés en profils qualitativement différents.
Compte tenu de la ségrégation des réseaux corticaux en processus descendant et ascendant, une prédiction évidente est que les dommages aux parties supérieures du lobule pariétal (SPL subsumante) devraient entraîner un déficit d’orientation attentionnelle dirigée vers l’objectif, tandis que les dommages aux parties inférieures du lobule pariétal (TPJ subsumante) entraîneraient un déficit associé à la capture de l’attention par stimulus. Dans la mesure où ces correspondances cerveau-comportement ont été explorées dans la littérature neuropsychologique, cette prédiction n’est évidemment pas confirmée. Par exemple, les symptômes cliniques de négligence hémispatiale sont fortement associés à des dommages aux parties inférieures du lobe pariétal, qui comprend la TPJ, plutôt qu’à des parties supérieures comme la SPL (Friedrich et al., 1998; Shomstein et coll., 2010; Corbetta et Shulman, 2011). Ceci est quelque peu en contradiction avec la littérature de neuroimagerie, qui suggère que le rôle de la TPJ est dans la capture de l’attention, plutôt que dans l’orientation volontaire de l’attention, le domaine dans lequel les patients négligents semblent avoir le plus de difficultés. Pour compliquer davantage les choses, il a été noté que les lésions qui impliquent exclusivement la LPS ne produisent que rarement des preuves cliniques de négligence (Vallar et Perani, 1986). Une autre étude récente avec des patients présentant des lésions centrées principalement sur la TPJ et la STG mais préservées SPL, Corbetta et al. (2005) ont montré que la négligence spatiale, ainsi que sa récupération, était associée à la restauration de l’activité dans les régions temporo-pariétale ventrale et pariétale dorsale (voir Corbetta et Shulman, 2011 pour une revue). Bien qu’intéressante et passionnante dans ses conclusions, cette dernière étude ne différencie pas la contribution relative des voies dorsales et ventrales aux différents types d’attention, puisque les patients n’ont été testés que sur une variante de la tâche de recherche d’attention spatiale cachée de Posner, tâche qui est censée engager à la fois une orientation attentionnelle descendante et ascendante.
Pour faire la distinction entre le contrôle attentionnel axé sur les objectifs et la capture attentionnelle saillante et pour examiner leur cartographie sur la SPL et la TPJ, respectivement, une étude récente a adopté deux paradigmes comportementaux, chacun ciblant l’une de ces formes d’attention (Shomstein et al., 2010). Pour examiner l’intégrité de l’orientation attentionnelle descendante chez les patients, une tâche descendante a été utilisée exigeant que les participants déplacent l’attention spatiale entre les flux RSVP séparés spatialement (une tâche qui a été utilisée avec succès pour démontrer l’activation de la SPL dans les études IRMF (Yantis et al., 2002)). De même, afin d’examiner les capacités d’orientation attentionnelle ascendantes des patients, une variante de Folk et al. (2002) ont utilisé un paradigme de capture contingente dans lequel les participants détectaient des cibles qui apparaissaient lors de la fixation tandis que des singletons de couleur non pertinents pour les tâches étaient flashés à la périphérie. La mesure dans laquelle les distracteurs non pertinents à la tâche interfèrent avec la tâche de détection centrale a ensuite été utilisée comme mesure de la capture attentionnelle ascendante (Bacon et Egeth, 1994; Folk et al., 2002).
Les prédictions étaient les suivantes: les patients présentant des lésions sur les parties supérieures du lobe pariétal (affectant la LPS) doivent être altérés dans la tâche d’orientation attentionnelle descendante (avec des performances préservées sur la tâche de capture) tandis que les patients présentant des lésions sur les parties inférieures du lobe pariétal (affectant la TPJ) doivent être altérés sur la tâche de capture (avec des performances épargnées sur la tâche descendante). Une double dissociation de cette forme atteste non seulement des composantes indépendantes de l’attention, mais suggère également que de telles composantes attentionnelles sont médiées par des mécanismes neuronaux indépendants. Huit patients atteints de négligence visuo-spatiale ont été recrutés pour l’étude et ont accompli deux tâches, en utilisant une orientation attentionnelle axée sur le stimulus ou dirigée vers un objectif. En fonction de leur profil comportemental, les patients ont été classés en groupes et le chevauchement de leurs lésions a été exploré (figure 2A). Les patients qui présentaient des difficultés d’orientation attentionnelle dirigée vers l’objectif, telles que quantifiées par l’indice attentionnel descendant (Figure 2B), présentaient un chevauchement de lésion centré sur des parties supérieures du lobule pariétal (SPL subsumante) avec un lobule pariétal inférieur épargné (TPJ). Les patients présentant un chevauchement de lésion centré sur les parties inférieures du lobule pariétal (TPJ subsumante) mais épargnés de SPL ont effectué normalement la tâche d’orientation dirigée vers l’objectif, tout en restant immunisés contre la capture attentionnelle (Figure 2C). Les résultats de cette étude suggèrent clairement que la SPL et la TPJ sont des régions anatomiques qui sont nécessairement recrutées à des fins d’orientation descendante et ascendante et que les dommages causés à la SPL et à la TPJ entraînent des troubles de l’orientation descendante et ascendante respectivement.
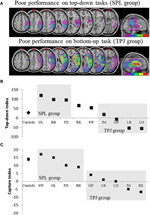
Figure 2. Résultats de l’étude neuropsychologique visant à étudier la contribution relative de la LPS et de la TPJ à l’orientation descendante et ascendante. (A) Chevauchements de lésions (chevauchement minimal violet; chevauchement maximal rouge) pour les patients regroupés par déficits comportementaux dans l’orientation attentionnelle descendante, étiquetés groupe SPL (panneau supérieur); et les patients regroupés par déficits comportementaux dans l’orientation ascendante, étiquetés groupe TPJ (panneau inférieur). (B) Performance comportementale sur la tâche descendante résumée par un « Indice descendant” qui quantifie les différences entre les déplacements spatiaux descendants effectués de gauche à droite et vice versa. Les témoins et le groupe lésé par la TPJ montrent des efficacités similaires dans l’exécution des déplacements spatiaux, tandis que les patients atteints de lésions SPL montrent une efficacité réduite. Les données de contrôle de groupe et de patient individuel (étiquetées avec les initiales du patient) sont tracées en abscisse. C) L’” indice de capture » est une mesure de l’attention ascendante et quantifie la mesure dans laquelle les distracteurs non pertinents de la tâche captent l’attention loin de la tâche. Les contrôles et le groupe mis en place par SPL affichent des valeurs de capture similaires, de sorte que les deux groupes sont capturés par les distracteurs non pertinents de la tâche. Le groupe mis en place par TPJ montre un indice de capture très réduit (échec de la capture). Notez que les patients ont été placés dans le groupe SPL ou TPJ en fonction du comportement, plutôt que de la lésion, notez donc la cohérence avec laquelle les patients se retrouvent dans le groupe correspondant.
Interaction Entre la sélection Descendante et ascendante
Bien qu’il existe apparemment une forte association entre l’orientation dirigée vers l’objectif et la SPL et l’orientation pilotée par le stimulus et la TPJ, les données de Shomstein et al. (2010) une étude de patients suggère que ces deux systèmes ne sont pas entièrement indépendants. Cette conclusion est étayée par la constatation que les patients présentant des lésions SPL présentaient un modèle de performance étiqueté comme « hyper capture. »Contrairement aux contrôles, pour lesquels seul le distracteur de couleur cible captait l’attention (entraînant une précision de cible inférieure), les distracteurs de couleur non pertinents se sont également révélés distrayants pour les patients atteints de lésion SPL. De plus, alors que pour les témoins, l’attention n’était captée par les distracteurs que lorsqu’ils précédaient l’apparition de la cible, pour les patients présentant des lésions SPL, l’attention était même captée par les distracteurs présentés simultanément avec la cible. Ce modèle de performance peut être expliqué par le cadre suivant: SPL est responsable de l’orientation descendante de l’attention qui comprend la détermination des aspects des stimuli qui sont pertinents pour la tâche (par exemple, rechercher une cible rouge; Corbetta et Shulman, 2002; Serences et al., 2005). Cet ensemble attentionnel contraint alors TPJ, de sorte que le mécanisme de capture de l’attention médiatisé par TPJ n’est déclenché que par les informations pertinentes de la tâche (par exemple, les distracteurs rouges capturant l’attention et les distracteurs gris ne capturant pas l’attention lors de la recherche d’une cible rouge). L’absence de SPL empêche l’établissement d’un ensemble attentionnel pertinent à la tâche et, par conséquent, tout stimulus, pertinent ou non à la tâche, est jugé important, capturant donc l’attention (par exemple, un distracteur non pertinent à la tâche capturant l’attention pour le groupe SPL) sans discernement.
Il a été suggéré que SPL et TPJ pourraient interagir d’au moins l’une des deux manières possibles. La première possibilité est que TPJ sert de système d’alerte qui détecte des stimuli pertinents sur le plan comportemental mais qui n’ont pas la haute résolution spatiale, ainsi lorsqu’un stimulus pertinent sur le plan comportemental est détecté, sa localisation précise est fournie par le SPL qui stocke des cartes spatiales (Kastner et al., 1999; Wojciulik et Kanwisher, 1999; Bisley et Goldberg, 2003; Silver et al., 2005). Une possibilité hypothétique connexe est que le mécanisme de capture (qui inclut la TPJ) agit comme un disjoncteur de l’activité cognitive en cours lorsqu’un stimulus pertinent sur le plan comportemental est présenté (Corbetta et Shulman, 2002, 2011). Le schéma d’activité « hyper-capture » observé chez les patients présentant une TPJ préservée mais une SPL lésée fournit une preuve supplémentaire de l’hypothèse selon laquelle la TPJ émet un signal de commande qui met fin à la tâche à accomplir, servant ainsi de disjoncteur (Corbetta et Shulman, 2002; Serences et al., 2005). D’autres études récentes de neuroimagerie utilisant divers paradigmes ont fourni des preuves supplémentaires d’une relation interactive entre l’orientation attentionnelle descendante et ascendante, puis de la relation entre SPL et TPJ (Serences et al., 2005; Asplund et coll., 2010; Diquattro et Geng, 2011).
Bien que les preuves d’une interaction entre les deux systèmes attentionnels et les deux substrats attentionnels (SPL et TPJ) soient solides, ce qui reste flou est de savoir si cette interaction est directe entre SPL et TPJ ou si elle est réalisée par d’autres régions intermédiaires. Comme mentionné précédemment, les réseaux d’orientation attentionnelle de haut en bas et de bas en haut engagent diverses régions du cortex frontal, il est donc raisonnable d’émettre l’hypothèse que la convergence entre les deux systèmes pourrait être réalisée via le lobe frontal. Deux études récentes portant sur l’interaction entre la sélection attentionnelle descendante et ascendante ont fourni des preuves pour la FIJ et l’IFG en tant que sites possibles de convergence entre la sélection axée sur le stimulus et la sélection dirigée vers des objectifs (Asplund et al., 2010; Diquattro et Geng, 2011). La FIJ et l’IFG semblent être les candidats idéaux pour une telle interaction compte tenu de leur implication générale dans l’attention et le contrôle cognitif ainsi que de leur implication dans la sélection spatiale et non spatiale (Koechlin et al., 2003; Brass et coll., 2005).
Le rôle du Lobe pariétal dans la Sélection de Haut en Bas et de Bas en Haut: Physiologie
Bien que l’accent de cette revue ait été principalement mis sur les études humaines, une grande richesse de connaissances sur l’implication du cortex pariétal dans l’orientation attentionnelle a été glanée à partir des recherches sur la physiologie du singe (voir revue récente de Bisley et Goldberg, 2010). Cependant, lorsqu’il s’agit d’examiner les contributions relatives de différentes régions du cortex pariétal à l’orientation attentionnelle descendante et ascendante, la littérature sur la physiologie du singe manque. La principale raison en est que dans le cortex de singe, il ne semble pas y avoir de preuve de la même ségrégation du contrôle descendant et ascendant. Au lieu de cela, la zone intrapariétale latérale (LÈVRE) initialement considérée comme impliquée dans la planification des saccades (Gnadt et Andersen, 1988) est impliquée dans l’attention visuelle et agit comme une carte de priorité dans laquelle les stimuli externes sont représentés en fonction de leur priorité comportementale dérivée de manière descendante ou ascendante (Colby et Goldberg, 1999; Bisley et Goldberg, 2003, 2010; Balan et Gottlieb, 2006; Ipata et al., 2006; Buschman et Miller, 2007; Gottlieb et Balan, 2010).
Conclusion
Bien que l’on en sache beaucoup moins sur le cortex pariétal humain que sur celui du cortex homologue de singe, des études récentes, utilisant des méthodes de neuroimagerie et de neuropsychologie, ont commencé à élucider des distinctions fonctionnelles et structurelles de plus en plus fines. Cette revue s’est concentrée sur des études récentes de neuroimagerie et de neuropsychologie élucidant les rôles cognitifs des régions dorsales et ventrales du cortex pariétal dans l’orientation attentionnelle de haut en bas et de bas en haut, et sur l’interaction entre les deux mécanismes d’allocation attentionnelle.
Déclaration de conflit d’intérêts
L’auteur déclare que la recherche a été menée en l’absence de relations commerciales ou financières pouvant être interprétées comme un conflit d’intérêts potentiel.
Remerciements
Ce travail a été soutenu par la subvention des National Institutes of Health EY021644.
Asplund, C. L., Todd, J. J., Snyder, A. P., et Marois, R. (2010). Un rôle central pour le cortex préfrontal latéral dans l’attention dirigée vers un objectif et dirigée par un stimulus. NAT. Neurosci. 13, 507–512.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Bacon, W. F., et Egeth, H.E. (1994). Capture attentionnelle axée sur le stimulus. Percept. Psychophiles. 55, 485–496.
Résumé Pubmed / Texte intégral Pubmed
Balan, P. F., et Gottlieb, J. (2006). Intégration d’intrants exogènes dans une carte de saillance dynamique révélée par l’attention perturbatrice. J. Neurosci. 26, 9239–9249.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Behrmann, M., Geng, J. J. et Shomstein, S. (2004). Cortex pariétal et attention. Curr. Opin. Neurobiol. 14, 212–217.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Bisley, J. W., et Goldberg, M. E. (2003). Activité neuronale dans la zone intrapariétale latérale et attention spatiale. Science 299, 81-86.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Bisley, J. W., et Goldberg, M. E. (2010). Attention, intention et priorité dans le lobe pariétal. Annu. Rév. Neurosci. 33, 1–21.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Brass, M., Derrfuss, J., Forstmann, B., et von Cramon, D. Y. (2005). Le rôle de la zone de jonction frontale inférieure dans le contrôle cognitif. Tendances Cogn. Sci. 9, 314–316.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Buschman, T. J., et Miller, E. K. (2007). Contrôle top-down versus bottom-up de l’attention dans les cortex pariétaux préfrontal et postérieur. Science 315, 1860-1862.
Résumé Pubmed | Texte intégral Pubmed / Texte intégral CrossRef
Chiu, Y. C., et Yantis, S. (2009). Une source de contrôle cognitif indépendante du domaine pour les ensembles de tâches: déplacer l’attention spatiale et changer de règles de catégorisation. J. Neurosci. 29, 3930–3938.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Colby, C. L., et Goldberg, M. E. (1999). Espace et attention dans le cortex pariétal. Annu. Rév. Neurosci. 22, 319–349.
Résumé Pubmed | Texte intégral Pubmed / Texte intégral Croisé
Corbetta, M., Kincade, J. M., Ollinger, J. M., McAvoy, M. P. et Shulman, G. L. (2000). L’orientation volontaire est dissociée de la détection de cible dans le cortex pariétal postérieur humain. NAT. Neurosci. 3, 292–297.
Résumé Pubmed | Texte intégral Pubmed / Texte intégral CrossRef
Corbetta, M., Kincade, M. J., Lewis, C., Snyder, A. Z., et Sapir, A. (2005). Base neurale et récupération des déficits d’attention spatiale dans la négligence spatiale. NAT. Neurosci. 8, 1603–1610.
Résumé Pubmed | Texte intégral Pubmed / Texte intégral CrossRef
Corbetta, M., Patel, G. et Shulman, G. L. (2008). Le système de réorientation du cerveau humain: de l’environnement à la théorie de l’esprit. Neurone 58, 306-324.
Résumé Pubmed | Texte intégral Pubmed / Texte intégral CrossRef
Corbetta, M., et Shulman, G. L. (2002). Contrôle de l’attention ciblée et stimulée par le stimulus dans le cerveau. NAT. Rév. Neurosci. 3, 201–215.
Résumé Pubmed | Texte intégral Pubmed / Texte intégral CrossRef
Corbetta, M., et Shulman, G. L. (2011). Négligence spatiale et réseaux d’attention. Annu. Rév. Neurosci. 34, 569–599.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Critchley, M. (1953). Les Lobes Pariétaux. Londres, Royaume-Uni: Hafner Press.
Résumé de Pubmed / Texte intégral de Pubmed
Diquattro, N.E., et Geng, J. J. (2011). La connaissance contextuelle configure les réseaux de contrôle attentionnel. J. Neurosci. 31, 18026–18035.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Downar, J., Crawley, A. P., Mikulis, D. J., et Davis, K. D. (2001). L’effet de la pertinence de la tâche sur la réponse corticale aux changements de stimuli visuels et auditifs: une étude IRMf liée aux événements. Neuroimage 14, 1256-1267.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Downar, J., Crawley, A. P., Mikulis, D. J., et Davis, K. D. (2002). Un réseau cortical sensible à la saillance du stimulus dans un contexte comportemental neutre à travers de multiples modalités sensorielles. J. Neurophysiol. 87, 615–620.
Résumé de Pubmed / Texte intégral de Pubmed
Drummond, L., et Shomstein, S. (2010). Attention basée sur les objets: changement ou incertitude? Attention. Percept. Psychophiles. 72, 1743–1755.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Egeth, H. E., et Yantis, S. (1997). Attention visuelle: contrôle, représentation et parcours temporel. Annu. Rév. Psychol. 48, 269–297.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Eriksen, C. W., et Hoffman, J.E. (1972). Caractéristiques temporelles et spatiales du codage sélectif à partir d’affichages visuels. Percept. Psychophiles. 12, 201–204.
Folk, C. L., Leber, A. B., et Egeth, H.E. (2002). Vous a fait cligner des yeux! La capture attentionnelle contingente produit un clignotement spatial. Percept. Psychophiles. 64, 741–753.
Résumé de Pubmed / Texte intégral de Pubmed
Folk, C. L., Remington, R. W. et Johnston, J.C. (1992). L’orientation secrète involontaire dépend des paramètres de contrôle de l’attention. J. Exp. Psychol. Hum. Percept. Effectuer. 18, 1030–1044.
Résumé de Pubmed / Texte intégral de Pubmed
Friedrich, F. J., Egly, R., Rafal, R. D., et Beck, D. (1998). Déficits d’attention spatiale chez l’homme: comparaison des lésions de jonction pariétale et temporo-pariétale supérieures. Neuropsychologie 12, 193-207.
Résumé de Pubmed / Texte intégral de Pubmed
Geng, J. J., et Behrmann, M. (2002). La recherche de probabilité de l’emplacement de la cible facilite implicitement la recherche visuelle chez les participants normaux et les patients souffrant de négligence hémispatiale. Psychol. sci. 13, 520–525.
Résumé de Pubmed / Texte intégral de Pubmed
Geng, J. J., et Behrmann, M. (2005). Probabilité spatiale comme repère attentionnel dans la recherche visuelle. Percept. Psychophiles. 67, 1252–1268.
Résumé de Pubmed / Texte intégral de Pubmed
Geng, J. J., et Mangun, G. R. (2011). L’activation de la jonction temporopariétale droite par un repère contextuel saillant facilite la discrimination de la cible. Neuroimage 54, 594-601.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Gibson, B. S., et Kelsey, E.M. (1998). La capture attentionnelle pilotée par le stimulus dépend de l’ensemble attentionnel pour les fonctionnalités visuelles à l’échelle de l’écran. J. Exp. Psychol. Hum. Percept. Effectuer. 24, 699–706.
Résumé de Pubmed / Texte intégral de Pubmed
Giesbrecht, B., Woldorff, M. G., Song, A. W., et Mangun, G. R. (2003). Mécanismes neuronaux de contrôle de haut en bas pendant l’attention spatiale et caractéristique. Neuroimage 19, 496-512.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Gnadt, J. W., et Andersen, R.A. (1988). Activité de planification motrice liée à la mémoire dans le cortex pariétal postérieur du macaque. Exp. Brain Res. 70, 216-220.
Résumé Pubmed / Texte intégral Pubmed
Gottlieb, J., et Balan, P. (2010). Attention comme décision dans l’espace d’information. Tendances Cogn. Sci. 14, 240–248.
Résumé Pubmed | Texte intégral Pubmed / Texte intégral CrossRef
Greenberg, A. S., Esterman, M., Wilson, D., Serences, J. T. et Yantis, S. (2010). Contrôle de l’attention spatiale et basée sur les caractéristiques dans le cortex frontopariétal. J. Neurosci. 30, 14330–14339.
Résumé Pubmed | Texte intégral Pubmed / Texte intégral CrossRef
Hopfinger, J. B., Buonocore, M. H. et Mangun, G. R. (2000). Les mécanismes neuronaux du contrôle attentionnel de haut en bas. NAT. Neurosci. 3, 284–291.
Résumé Pubmed / Texte Intégral Pubmed / Texte intégral CrossRef
Ipata, A. E., Gee, A. L., Gottlieb, J., Bisley, J. W. et Goldberg, M. E. (2006). Les réponses des lèvres à un stimulus popout sont réduites si elles sont ouvertement ignorées. NAT. Neurosci. 9, 1071–1076.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Johnston, W. A., et Dark, V. J. (1986). Attention sélective. Annu. Rév. Psychol. 37, 43–75.
Juola, J. F., Koshino, H. et Warner, C. B. (1995). Compromis entre les effets attentionnels des signaux spatiaux et les effets brusques. Percept. Psychophiles. 57, 333–342.
Résumé de Pubmed / Texte intégral de Pubmed
Kastner, S., Pinsk, M. A., De Weerd, P., Desimone, R., et Ungerleider, L. G. (1999). Activité accrue dans le cortex visuel humain pendant l’attention dirigée en l’absence de stimulation visuelle. Neurone 22, 751-761.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Koechlin, E., Ody, C. et Kouneiher, F. (2003). L’architecture du contrôle cognitif dans le cortex préfrontal humain. Science 302, 1181-1185.
Résumé Pubmed / Texte Intégral Pubmed / Texte intégral CrossRef
Koshino, H., Warner, C. B., et Juola, J. F. (1992). Efficacité relative des signaux centraux, périphériques et d’apparition brutale de l’attention visuelle. Q. J. Exp. Psychol. A 45, 609-631.
Résumé de Pubmed / Texte intégral de Pubmed
Liu, T., Slotnick, S. D., Serences, J. T. et Yantis, S. (2003). Mécanismes corticaux de contrôle attentionnel basé sur les caractéristiques. Céreb. Cortex 13, 1334-1343.
Résumé Pubmed | Texte intégral Pubmed / Texte intégral CrossRef
Marois, R., Leung, H. C., et Gore, J. C. (2000). Une approche axée sur le stimulus du traitement de l’identité et de la localisation des objets dans le cerveau humain. Neurone 25, 717-728.
Résumé Pubmed | Texte intégral Pubmed / Texte intégral CrossRef
McFire, J., et Zangwill, O. L. (1960). Handicaps visuo-constructifs associés à des lésions de l’hémisphère cérébral gauche. Cerveau 82, 243-260.
Moore, C. M., et Egeth, H. (1998). Comment l’attention basée sur les fonctionnalités affecte-t-elle le traitement visuel? J. Exp. Psychol. Hum. Percept. Effectuer. 24, 1296–1310.
Résumé de Pubmed / Texte intégral de Pubmed
Moran, J., et Desimone, R. (1985). L’attention sélective bloque le traitement visuel dans le cortex extrastrié. Science 229, 782-784.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
O’Craven, K., Rosen, B. R., Kwong, K. K., Treisman, A. et Savoy, R. L. (1997). L’attention volontaire module l’activité de l’IRMf chez l’homme MT-MST. Neurone 18, 591-598.
Résumé Pubmed | Texte intégral Pubmed / Texte intégral CrossRef
Piercy, M. (1964). Les effets des lésions cérébrales sur la fonction intellectuelle: une revue des tendances actuelles de la recherche. Fr. J. Psychiatrie 110, 310-352.
Résumé de Pubmed / Texte intégral de Pubmed
Posner, M. I. (1980). Orientation de l’attention. Q. J. Exp. Psychol. 32, 3–25.
Résumé de Pubmed / Texte intégral de Pubmed
Posner, M. I., Snyder, C. R., et Davidson, B. J. (1980). Attention et détection des signaux. J. Exp. Psychol. 109, 160–174.
Résumé de Pubmed / Texte intégral de Pubmed
Posner, M. I., Walker, J. A., Friedrich, F. J., et Rafal, R. D. (1984). Effets de la lésion pariétale sur l’orientation secrète de l’attention. J. Neurosci. 4, 1863–1874.
Résumé de Pubmed / Texte intégral de Pubmed
Serences, J. T., Shomstein, S., Leber, A. B., Golay, X., Egeth, H. E. et Yantis, S. (2005). Coordination du contrôle attentionnel volontaire et impulsé par le stimulus dans le cortex humain. Psychol. Sci. 16, 114–122.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Shaw, M. L. (1978). Un modèle d’allocation de capacité pour le temps de réaction. J. Exp. Psychol. Hum. Percept. Effectuer. 4, 586–598.
Shomstein, S., et Behrmann, M. (2006). Systèmes corticaux médiant l’attention visuelle à la fois sur les objets et les emplacements spatiaux. Proc. Natl. Acad. Sci. États-Unis 103, 11387-11392.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Shomstein, S., Lee, J. et Behrmann, M. (2010). Guidage attentionnel de haut en bas et de bas en haut: étude du rôle des cortex pariétaux dorsal et ventral. Exp. Brain Res. 206, 197-208.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Shomstein, S., et Yantis, S. (2004a). Hiérarchisation configurale et contextuelle dans l’attention basée sur les objets. Psychon. Taureau. Rév. 11, 247-253.
Résumé de Pubmed / Texte intégral de Pubmed
Shomstein, S., et Yantis, S. (2004b). Le contrôle de l’attention se déplace entre la vision et l’audition dans le cortex humain. J. Neurosci. 24, 10702–10706.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Shomstein, S., et Yantis, S. (2006). Le cortex pariétal médie le contrôle volontaire de l’attention auditive spatiale et non spatiale. J. Neurosci. 26, 435–439.
Résumé Pubmed | Texte Intégral Pubmed / Texte intégral CrossRef
Silver, M. A., Ress, D., et Heeger, D.J. (2005). Cartes topographiques de l’attention spatiale visuelle dans le cortex pariétal humain. J. Neurophysiol. 94, 1358–1371.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Tamber-Rosenau, B. J., Esterman, M., Chiu, Y.-C. et Yantis, S. (2011). Mécanismes corticaux de contrôle cognitif pour déplacer l’attention dans la vision et la mémoire de travail. J. Cogn. Neurosci. 23, 2905–2919.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Theeuwes, J. (1991). Contrôle exogène et endogène de l’attention: l’effet des effets visuels et des décalages. Percept. Psychophiles. 49, 83–90.
Résumé de Pubmed / Texte intégral de Pubmed
Vallar, G., et Perani, D. (1986). L’anatomie de la négligence unilatérale après des lésions d’AVC de l’hémisphère droit. Une étude de corrélation clinique / CT-scan chez l’homme. Neuropsychologie 24, 609-622.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Wojciulik, E., et Kanwisher, N. (1999). La généralité de l’implication pariétale dans l’attention visuelle. Neurone 23, 747-764.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Yantis, S., et Jonides, J. (1984). Effets visuels brusques et attention sélective: preuves de la recherche visuelle. J. Exp. Psychol. Hum. Percept. Effectuer. 10, 601–621.
Résumé de Pubmed / Texte intégral de Pubmed
Yantis, S., et Jonides, J. (1990). Effets visuels brusques et attention sélective: allocation volontaire ou automatique. J. Exp. Psychol. Hum. Percept. Effectuer. 16, 121–134.
Résumé de Pubmed / Texte intégral de Pubmed
Yantis, S., Schwarzbach, J., Serences, J. T. Il s’agit de l’une des principales sources d’information sur la santé et la sécurité au travail. Activité neuronale transitoire dans le cortex pariétal humain pendant les changements d’attention spatiale. NAT. Neurosci. 5, 995–1002.
Résumé Pubmed / Texte intégral Pubmed / Texte intégral CrossRef
Yantis, S., et Serences, J. T. (2003). Mécanismes corticaux de contrôle attentionnel basé sur l’espace et l’objet. Curr. Opin. Neurobiol. 13, 187–193.
Résumé Pubmed / Texte Intégral Pubmed / Texte intégral CrossRef