Invertebrate Anatomy OnLine
Daphnia magna ©
Water Flea
19jun2006
Copyright 2000 by
Richard Fox
Lander University
Preface
This is one of many exercises available from Invertebrate Anatomy OnLine , an Internet laboratory manual for courses in Invertebrate Zoology. 追加の演習は、左へのリンクをクリックしてアクセスすることができます。 供給および実験室の技術の用語集そして章はまた利用できる。 これらの演習で使用される用語と系統発生は、Ruppert、Fox、およびBarnes(2004)による無脊椎動物動物学の教科書での使用に対応しています。 ハイフネーションされた図の吹き出しは、教科書の図を参照してください。 ハイフネーションされていないコールアウトは、演習に埋め込まれた図を参照します。 用語集には、この教科書の用語だけでなく、実験室の演習が含まれています。
系統学
節足動物P
節足動物は、はるかに最大かつ最も多様な動物分類群では、chelicerates、昆虫、myriapods、甲殻類だけでなく、Trilobitomorphaなどの多くの絶滅 分節された体は、各分節に一対の接合された付属器を原始的に有する。 表皮は複雑なクチクラ外骨格を分泌し、サイズの増加を可能にするために脱皮しなければならない。 現存する節足動物は、セグメントと付属器の構造と機能に地域的な特殊化を示しますが、祖先はおそらくすべてのセグメントに同様の付属器を持っていました。 体は典型的には頭と胴に分かれており、その胴はしばしばさらに胸郭と腹部に分かれています。 腸は前腸、中腸、および後腸で構成され、前口から後肛門まで体の長さを延長します。
腸は前腸、中腸、および後腸で構成され、体の長さを延長します。
前腸および後腸は、それぞれ胚性stomodeumおよびproctodeumに由来する表皮陥入であり、節足動物のすべての表皮表面と同様に、クチクラによって裏打ちされている。 中腸は内胚葉であり、ほとんどの酵素分泌、加水分解および吸収を担う。体腔は生殖腺と腎臓に関連する小さな空間に縮小されます。
体腔は、生殖腺と腎臓に関連する小さな空間に縮小されます。
体腔 機能体腔は、水平横隔膜によって背側心膜洞およびはるかに大きな心膜洞に分割された広々としたヘモコエルである。 時には、腹側神経索を取り囲む小さな腹側神経周囲洞がある。 hemalシステムはhemocoelに血をポンプでくむ背側の、収縮の、管状の、ostiate中心を含んでいます。
hemalシステムはhemocoelに血をポンプでくむ背側の、収縮の、管状の、ostiate中心を 排泄器官は分類群によって異なり、Malpighian tubules、saccate nephridia、およびnephrocytesが含まれています。 呼吸器も分類群によって異なり、鰓、本肺、気管の多くの種類が含まれています。
神経系は、2つまたは3つの対の神経節、circumenteric connectives、および分節性神経節および分節性末梢神経を有する対の腹側神経コードの背側、前脳 様々な程度の凝縮およびcephalizationは、異なる分類群に見られる。
開発はcentrolecithal卵と表面的な切断で派生しています。 多くの場合、幼虫が発生しますが、多くの場合は直接発生します。 幼体は、成体の大きさと生殖状態に達するまで、脱皮によって分離された一連の甲殻類を通過する。 この時点で脱皮と成長は、分類群に応じて、停止または継続することができます。
Mandibulata
MandibulataはChelicerataの姉妹分類群であり、対照的に、最初の頭のセグメントに触角、第三に下顎、第四に上顎を持っています。 脳は三対の神経節を持つsyncerebrumであり、二つの神経節ではない。 祖先の下顎は、おそらくbiramous付属器とJ字型の腸、後向きの口、および腹側の食物溝を持っていました。 二つの最高レベルの下顎の分類群は、甲殻類と気管です。
甲殻類sP
甲殻類は気管の姉妹分類群であり、第二の頭部セグメントに触角を有する点が異なり、合計2対であり、これは一意である。 元の甲殻類の付属肢はbiramousであったが、uniramous肢は派生分類群で共通している。 元のタグマタは頭であったが、これは多くの分類群で頭、胸郭、腹部または頭胸郭と腹部に置き換えられている。 排泄は嚢状腎症の一つ、時には二つのペアを介して行われ、呼吸は、時には体表面によって、えらの多種多様によって達成されます。 ナウプリウスは最も初期の孵化段階であり、ナウプリウスの目は三つまたは四つの中央のocelliで構成されています。
Eucrustacea
Eucrustaceaには、レミペデスを除くすべての最近の甲殻類が含まれています。 分類群は熱、胸郭および腹部から成っている第一次tagmosisによってcephalothoraxおよび腹部の得られた状態がより共通であるが特徴付けられます。 八は胸部セグメントの最大数です。
胸脚
先祖の胸脚では、胸部付属器は、腹側の食物溝と一緒に懸濁給餌に使用される膨圧付属器であった。 このような付属肢と摂食は、いくつかの最近の分類群で持続するが、他の多くの分類群で修正されている。
Phyllopodomorpha
派生した固着目は多くの分類群で発生しますが、複眼は原始的にストーカーされています。
Phyllopoda
Phyllopodaは、”小さなphyllopodans”であるNotostraca、Laevicaudata、およびSpinicaudataおよびCladoceraからなる”大きなphyllopodans”の約800種で構成されています。 体幹の付属物は葉足類であり、大きな甲羅は体の大部分またはすべてを囲む。 大型のフィロポドン類は典型的には魚がいない遺物の習慣に生息するが、クラドケラン類はそのような制限を示さない。 Tagmataは頭部、胸郭および減らされた腹部です。 腹部は付属器を欠いているが、テルソン上に後部尾部のファーカを有する。 腹側の食物溝は通常存在し、摂食に使用される。 いわゆる背側器官は、後頭部の背側正中線上に存在する。
クラドセラO
クラドセラの11家族は、主に淡水プランクトンと底生甲殻類の約800種が含まれています。 クラドケラン、または水ノミは、小さな(0.2-6ミリメートル)水生甲殻類です。 ほとんどは静かな新鮮な水に生息しています。 ワタリムシやカイポッドと一緒に、彼らは淡水動物プランクトンのほとんどを占めています。 p>
ほとんどの分類群の甲羅は大きく、二枚貝であるため、頭を除くすべての体を囲みます。 二つの複眼は正中線上で融合している。 Nautliar目は人生を通して存在しています。 体は横方向に圧縮されている。 拡大した第二触角は歩行(水泳)器官であるが、第一触角は女性では痕跡的であり、男性ではあまり大きくない。 胸郭は短く、腹部には付属器がない。
ほとんどは、彼らが胸部付属器のセタを使用して水からフィルタリング植物プランクトンを消費する懸濁フィーダーです。 いくつかは、他のクラドセランを捕食肉食動物です。
クラドケランは単為生殖であり、年間の人口のほとんどは完全に無性生殖する女性で構成されています。 男性はめったに見られません。 甲羅は胚が保持され、直接開発が起こるひなの袋を囲みます。 冬(または時には夏)が近づくと、男性が現れ、有性生殖が起こり、耐性のある越冬卵が産生されます。 このような卵は、休息している卵が単為生殖の雌に孵化するまで、湖または池の底の堆積物に載っているpurselike ephippiumに囲まれていてもよい。
アノモポッドsO
アノモポッドは短い幹と大きな二枚貝の甲羅を持っています。 頭部は、頭部シールドを形成するために、背側および横方向に拡張される。 発達は直接的であり、休息している卵はephippiumに囲まれています。
実験室の標本
クラドケランの解剖学の研究は、地元の湖や池でプランクトンのトウで収集された生きた標本、またはCarolina BiologicalまたはWard Natural Scienceから入手可能な安価な培養物に基づくことができる。 Daphnia magnaはこの目的に適しています。 それはクラドケランが行くように、非常に大きな種であり、実験室の培養物で容易に維持されるものである。
外部解剖学
池の8センチ培養皿にいくつかのミジンコを配置し、解剖顕微鏡を使用して動物を観察します。 特徴的なぎくしゃくした水泳の動きに注意してください。 この動きの不均一な外観は、歩行付属器、またはオールの唯一のペアがあることの結果です。 水泳の付属物である大きな第二のアンテナの動きを観察してみてください。 観察は、動物が皿の底部に対して表面膜中に固定化されるように皿から水の大部分を除去することによって改善され得る。
これらの動物が可能である効果的な回避行動に注意して、大口径のプラスチック製のピペットで個体を捕獲する。 ワックスのフィートが付いているcoverslipを使用して単一の標本のぬれた台紙を準備しなさい。 これらの動物の厚さのために、彼らはカバースリップをサポートするために厚い足を必要とします。 スライドを複合顕微鏡のステージに置き、必要に応じて40Xと100Xで検査します。 特にそうするように言われなければ400Xを使用しないで下さい。
Tagmata
ミジンコの体は横方向に圧縮されており、カバースリップの圧力によって誇張された状態です。 体は前頭部、中胸部、および後腹部に分かれている(図1、19-15A)。 胸郭と腹部は甲羅に囲まれていますが、頭はそうではなく、甲羅の前に前方に伸びています。
頭
クラドケラン頭は腹側に曲がっており、ミジンコの頭は滑らかに丸みを帯びています。 多くの種では、頭の前端はプロセスまたは頭脊椎に生成される(図19-15B)が、それはD.magnaにはない。 ミジンコの頭部の腹側領域は、尖った吻部を形成するように拡張されている(図1)。 頭部は背側に体の残りの部分に接合されているが、深い裂け目によって腹側から分離されている。
大きなbiramous第二のアンテナは、頭の最も顕著な特徴であり、一つは、頭の中央付近の各側に発生します(図1)。 第二のアンテナは、強力な筋肉によって制御される単一の基底物品、花柄からなる。 筋肉は透明な体の壁を通して見えます。 二つのラミは、花柄の遠位端から生じる。 D.magnaでは、上のラムスに四つの記事があり、下のラムスには三つの記事があります。
二つのラミは大きな梅natatory setaeを負担します。 より高いパワーでsetaeを見て、大きなsetaeに小さな羽状に配置されたsetaeの行に注意してください。 結果は羽毛のようなものであり、したがって形容詞、羽毛、羽毛のようなものである。 触角はオールであり、セタエはパワーストローク中に水と接触する表面積を増加させる。 回復打撃の間にそれらは倒れ、表面積は減る。
女性の最初のアンテナは非常に小さいですが、おそらく演壇の先端の後ろにあるだけで見ることができます(図1)。 それは感覚機能を持っている男性で大きいですが、あなたの標本が男性であることは非常に低いです。
下顎骨はよく発達していますが、全量では容易には明らかではありません。 それぞれは、遠位端に硬化した歯を有する長い楕円形である(図1、19-15A)。 左右の下顎骨の歯は、頭の後端の口の両側にあります。p>
最初の上顎は非常に小さく、第二の上顎は存在しません。
祖先(および胚)の対になった側方複眼は、成人の正中線上で融合され、頭の内部に単一の中央複眼を形成する。 を搭載しているので、動きが見えてくるかもしれませんね。 筋肉は目から後方に伸びる。 また、目の内部の黒い色素の周囲に小さなレンズの集まりを見ることもできます。 P>
D.magnaとDを含む多くのクラドケラン、。 pulexは、単一の、小さな、中央値、nautpliar目、またはocellus、はるかに大きな複眼に後部および腹側を持っています。 それは脳の端に埋め込まれており、慎重な光調整と焦点合わせで目に見えるかもしれません(図1)。
図1ミジンコの雌。 Freeman&Bracegirdle(1971)から再描画されます。 Clad99L.gif p>
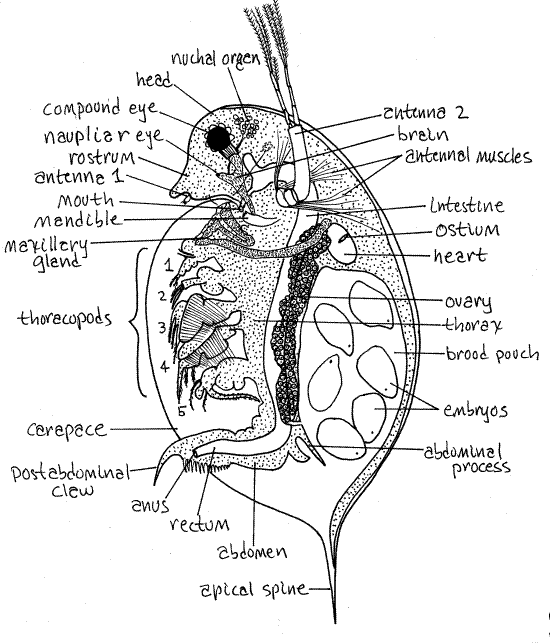
口は腹側の頭に位置し、後方を指していますが、おそらく表示されません。 それは2つの下顎骨の間にあります。
いくつかの種では、背正中線上の浅いノッチ、頸部洞が頭を胸郭から分離する。 ミジンコには子宮頸部洞がありません。 いくつかの種は、頸部洞の直後の正中線の外皮に背側器官を有するが、ミジンコはそうではない。
甲羅
甲羅は、posteriormostヘッドセグメントからposteriormostと横方向に延びる体壁の巨大な二重の折り目です。 それは首の後ろにのみ取り付けられた岬と比較されています。 ほとんどのcladocerans(anomopodsおよびctenopods)では、背側の正中線に沿って折り畳まれた大きくて薄くて柔軟なシートが2つの弁を形成し、動物の両側に1つがあります。 横方向に圧縮されています。 少数のonychopodおよびhaplopod cladoceransでは、それは比較的小さなひなの袋に還元され、幹を囲まない(図19-16A、B)。 甲羅は背側に閉じられているが、腹側に閉じられ、その二つの弁は、摂食電流の出入りを可能にするために裂けている(図19-15B)。
ミジンコを含む多くのクラドケランは、その機能が捕食を妨害する可能性のある後部頂端脊椎(=甲羅脊椎)を有する(図1、19-17B)。 多くの種はまた、頭の上に背骨を持ち、時には非常に大きなものを持っています。 プランクトン性クラドケランは、動物プランクトン性の魚や他の無脊椎動物、特にファントムミッジ、Chaoborusの幼虫によって消費されます。
甲羅は透明で、通常は無色または黄色です。 内部器官の大部分、および胸部付属器は、その透明な壁を通して見える。
胸郭
胸郭は頭のすぐ後ろにあり、体の残りの部分のほとんどを占めています。 それは完全に甲羅に囲まれています。
ミジンコの胸郭は、甲羅の腹側の裂け目から伸びる可能性のある遠位の先端が5対のbiramous、setose thoracopods(=胸部付属器)を負います(図1)。 これらの付属物の4つは供給の流れを発生させます。 彼らの動きは、腹側正中線からsetalフィルターを通って横方向に水を引き、そこから横方向に出る手足の間の空間に水を引きます(図19-12)。 食物粒子は、setalフィルターによって停止され、腹側の食物溝に保持され、腹側の食物溝では、それらが後方に指向された口に到達するまで、それらが付属器か
>1a.乾燥した培養皿の中心にワセリンの小さな軽打を置きます。 大口径のピペットでクラドセランを捕獲し、ゼリーの横に置きます。 それを傷つけないでゼリーに逆さまにcladoceranを押すのにあなたの良い鉗子を使用して下さい。 鉗子で絞ってはいけません。 彼らは圧迫されていることの絶対に不寛容であり、それはそれらを殺すでしょう。 あなたが成功する前に、いくつかの動物でこれを試してみる必要があります。
ゼリーにクラドセランを確保したら、動物が浸漬されるまで慎重に湖の水を加えます。 解剖顕微鏡の40倍で逆さまの生き物を観察する(図19-15B)。 付属器の動きに特に注意してください。 第二のアンテナを見てください。 甲羅の腹側の裂け目を見て、胸脚を見てください。 腹部の動きを観察する。 水の中にカルミン懸濁液の小さな滴を置き、水泳や摂食の流れの中の粒子の流れパターンを見てください。 <
腹部
あなたの全量の研究を再開します。 後方に胸郭が狭くなり、腹部になる(図1)。 腹部は付属器を欠いていますが、柔軟性があり、筋肉があり、非常に可動性があります。 それは通常、胸郭の下に押し込むために鋭く前方に曲がっており、そこで甲羅に完全に囲まれています。
肛門は腹部の後端で開きます。 腹後爪のペアは、腹部から後方に延びています。 腹部およびその爪は、胸部付属器を洗浄し、糸状藻類などの閉塞をフィルタ供給装置に除去するために使用される。 腹部の背側縁から長い羽毛の腹部のセットのペアが延びています。
二つの長い腹部プロセスは、腹部の背側縁から背側に延びています。 それらはひなの袋を閉め、卵または陰気な少年の早すぎる解放を防ぐためにドアとして作用する。 女性は彼女が部屋から彼女のひなを追い出すことを望むとき腹部プロセスを下げる。
腹部をまっすぐにして、甲羅から後方に伸ばすことができます。 動物はそれがこれを行うときに蹴っているように見え、あなたの標本が生きているならば、あなたは間違いなくそれがこれをするのを見るでしょう。
ヘマルシステム
ヘマルシステムは、心臓、hemocoel、および血液で構成されています。 ヘモコエルは体腔である。 短い楕円形の心臓は、前胸郭の背側領域の顕著な特徴である(図1、19-15A)。 心臓はhemocoelに囲まれています。p>
心臓はオスティアの単一のペアを持っていますが、これらは明らかではないかもしれません。 血管は存在しない。 心臓の収縮は、血液を前方に頭部のヘモコエルに押し込み、そこから3つのヘモコエルチャネルを介して胸部に後方に流れる。 中央チャネルは腸に腹側を実行し、胸部付属器に枝を放つのに対し、二つの側チャネルは、それぞれ甲羅の片側を提供しています。 甲羅は主なガス交換面である。 血液は、これらの領域のそれぞれから心臓に戻ります。
消化器系
C字型の腸は、少なくともその長さの一部については、口から背中の胸郭を通って、腹部を通って腹部の遠位端近くの肛門まで延びている(図1、19-15A)。 食物で満たされた腸の領域は容易に見られ、植物プランクトンで緑色であるか、または乾燥したクラドセランペレットで茶色である可能性が高い。 空の領域はより見えにくくなります。 短い憩室のペア、消化管は、前中腸から発生し、頭の中に延びています(図1、19-15A)
生殖器系
生殖腺は、腸の両側の胸郭の長さの大部分を延 Gonoductsは胸部の付属物の最後の組に後部のgonoporesによって外面に開く。 雌では、卵巣は卵管を介して背側に開き、雛室に入る。 男性では、精管は、腹腔後の腹側のgonoporeにつながる。
女性のクラドケランは、背甲の後方に位置する大きな水で満たされたひな袋を持っています(図1、19-15A)。 卵は卵管からここに押し出され、胚発生を完了して若年性クラドケランになるまでブロードされる。 この時点で、彼らは解放され、独立した存在を開始します。 あなたの標本のひなの袋は、卵、胚、または少年で満たされていてもよいです。 他の初期の生活史の段階を見るためにあなたのクラスメートの標本をどれを見ているかを判断してみてください。
女性は二つのタイプの卵を生産します。 夏の卵は卵黄がほとんどなく、受精せずに単為生殖的に発達する。 夏の卵は、通常、多くの、彼らは性的に成熟し、自分の若いを持っているまで、彼らは孵化するまで、いくつかの種では、少なくともひなチャンバーで運ばれます。 一方、休息している卵は非常に卵黄で厚い殻を持ち、受精後にのみ生産されます。
休息している卵は非常に卵黄で厚い殻を持ち、受精後 残りの卵は2個のみで、それぞれの卵巣から1個が生産されています。 これらの卵はまた、ひな室に放出されるが、夏の卵がなかったのに対し、受精されます。 冬の卵は、夏の卵とは異なり、むしろ雌によって迅速に放出され、分類群に応じて保護クチクラエフィピウムに囲まれているか、裸である。 雌が脱皮するとエフィピウムが放出される。 エフィピアは、種によって、沈むか、または浮かぶかもしれません。 冬の卵は翌年の春に孵化する。 冬の卵は常に単為生殖の雌、すなわち受精せずに繁殖する雌に孵化する。 最終的には、単為生殖の雌とその夏の卵の1つまたは数世代の後、男性が生産され、受精が行われて新しい世代の休息卵が生成されます。
>1b.利用可能な場合は、ミジンコエフィッピアの全量スライドを見てください(図19-17A)。 それぞれの2つの卵は、適切な照明で見えるはずです。 <
淡水プランクトン
>1c。 地元の湖や池からの新鮮なプランクトンのコレクションは、研究室で利用可能な場合があります。 もしそうなら、あなたの解剖顕微鏡にそれの小さな皿を取り、それを調べます。 あなたが存在する種のいくつかを区別できるかどうかを確認してください。 異なった種の個人を捕獲し、混合の顕微鏡との検査のためのそれらのwetmountsを作るのに大きい穴のプラスチックピペットを使用して下さい。 コレクションには、カイポッドと様々なワムシも含まれます。 幼虫はクラドケラン類やカイツブリ類よりもはるかに小さく、表面的には繊毛虫の原生動物に似ています。 Copepod naupliiもおそらく存在するでしょう(図19-8)。 <
ドッドソンSI、フレイDG。 1991. ThorpのCladoceraおよび他のBranchiopoda、J.H.&A.P.Covich(eds)。 北米の淡水無脊椎動物の生態学と分類。 学術出版社、サンディエゴ。
フリーマンWH,Bracegirdle B.1971. 無脊椎動物の構造のアトラス。 ヒエネマン教育の本、ロンドン。 129頁p>
ペナックRW。 1989. 米国の淡水無脊椎動物、3rd ed。 ニューヨーク州ワイリー出身。
Ruppert EE、Fox RS、Barnes RB。 2004. 無脊椎動物動物学、機能的進化的アプローチ、7th ed。 ブルックス-コール-トムソン、ベルモント-カリフォルニア州。 963頁
供給
大ボアプラスチックパストゥールピペット(標準的なプラスチックパストゥールピペットの先端をカット)
6-cm培養皿
湖の水
湖の水
湖の水
湖の水
湖の水
/p>
蜜蝋、センチメートルルール付き解剖キット
解剖顕微鏡
複合顕微鏡
ワセリン
ワセリン
ワセリン
ミジンコの文化
エフィピアの全山スライド
カルミン 懸濁液