Læringsmål
ved slutten av denne delen vil du kunne:
- Forklar prosessene for glykolyse
- Beskriv veien til et pyruvatmolekyl gjennom Krebs-syklusen
- Forklar transport av elektroner gjennom elektrontransportkjeden
- Beskriv PROSESSEN MED ATP-produksjon gjennom oksidativ fosforylering
- Oppsummere prosessen med glukoneogenese
Karbohydrater Er organiske molekyler som består av karbon, hydrogen og oksygenatomer. Familien av karbohydrater inneholder både enkle og komplekse sukkerarter. Glukose og fruktose er eksempler på enkle sukkerarter, og stivelse, glykogen og cellulose er alle eksempler på komplekse sukkerarter. De komplekse sukkerene kalles også polysakkarider og er laget av flere monosakkaridmolekyler. Polysakkarider tjener som energilagring (f.eks. stivelse og glykogen) og som strukturelle komponenter (f. eks. kitin i insekter og cellulose i planter).under fordøyelsen brytes karbohydrater ned i enkle, oppløselige sukkerarter som kan transporteres over tarmveggen inn i sirkulasjonssystemet som skal transporteres gjennom hele kroppen. Karbohydratfordøyelsen begynner i munnen med virkningen av spyttamylase på stivelse og slutter med monosakkarider som absorberes over tynntarmens epitel. Når de absorberte monosakkaridene transporteres til vevet, begynner prosessen med cellulær respirasjon (Figur 1). Denne delen vil først fokusere på glykolyse, en prosess hvor monosakkaridglukosen oksyderes, og frigjør energien som er lagret i bindingene for å produsere ATP.
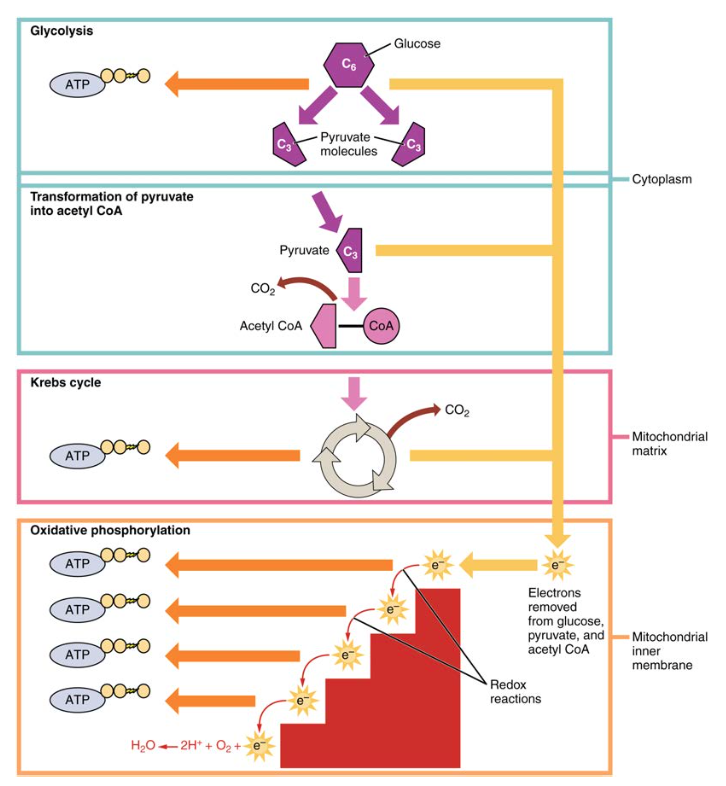
Figur 1. Cellulær respirasjon oksiderer glukosemolekyler gjennom glykolyse, Krebs-syklusen og oksidativ fosforylering for å produsere ATP.
Glykolyse
Glukose Er kroppens lettest tilgjengelige energikilde. Etter fordøyelsesprosesser bryter polysakkarider ned i monosakkarider, inkludert glukose, transporteres monosakkaridene over tynntarmens vegg og inn i sirkulasjonssystemet, som transporterer dem til leveren. I leveren passerer hepatocytter enten glukosen gjennom sirkulasjonssystemet eller lagrer overflødig glukose som glykogen. Celler i kroppen tar opp sirkulerende glukose som respons på insulin og, gjennom en rekke reaksjoner kalt glykolyse, overfører noe av energien i glukose TIL ADP for å danne ATP (Figur 2). Det siste trinnet i glykolyse produserer produktet pyruvat.
Glykolyse begynner med fosforylering av glukose ved heksokinase for å danne glukose-6-fosfat. Dette trinnet bruker EN ATP, som er donor av fosfatgruppen. Under virkningen av fosfofructokinase omdannes glukose-6-fosfat til fruktose-6-fosfat. På dette punktet donerer EN ANDRE ATP sin fosfatgruppe, som danner fruktose-1,6-bisfosfat. Dette seks-karbon sukker er delt for å danne to fosforylerte tre-karbon molekyler, glyceraldehyd-3-fosfat og dihydroksyaceton fosfat, som begge omdannes til glyceraldehyd-3-fosfat. Glyceraldehyd-3-fosfat fosforyleres videre med grupper donert av dihydrogenfosfat tilstede i cellen for å danne trekarbonmolekylet 1,3-bisfosfoglyserat. Energien til denne reaksjonen kommer fra oksidasjon av (fjerning av elektroner fra) glyceraldehyd-3-fosfat. I en rekke reaksjoner som fører til pyruvat, blir de to fosfatgruppene deretter overført til to Adp-er for å danne to Atp-er. Således bruker glykolyse to Atp-er, men genererer fire Atp-er, noe som gir en nettogevinst på to Atp-er og to pyruvatmolekyler. I nærvær av oksygen fortsetter pyruvat videre Til Krebs-syklusen (også kalt sitronsyresyklusen eller trikarboksylsyresyklusen (tca), hvor ytterligere energi ekstraheres og videreføres.
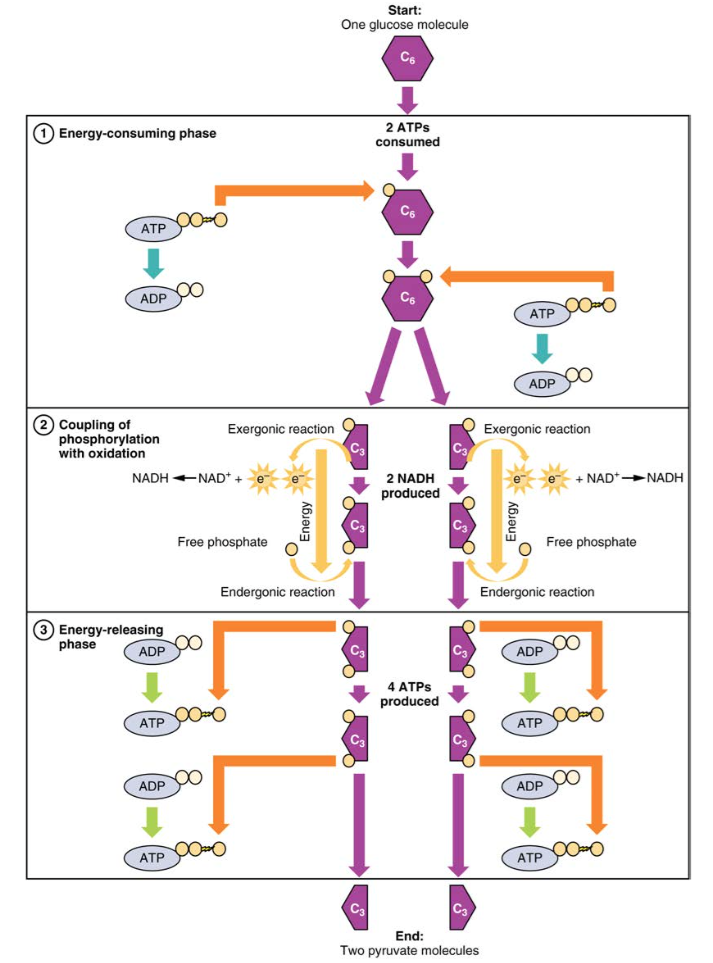
Figur 2. Under den energikrevende fasen av glykolyse forbrukes to Atp, og overfører to fosfater til glukosemolekylet. Glukosemolekylet splittes deretter i to tre-karbonforbindelser, som hver inneholder et fosfat. I løpet av den andre fasen tilsettes et ekstra fosfat til hver av de tre karbonforbindelsene. Energien til denne endergoniske reaksjonen er gitt ved fjerning (oksidasjon) av to elektroner fra hver tre-karbonforbindelse. Under energifrigjøringsfasen fjernes fosfatene fra begge tre karbonforbindelser og brukes til å produsere fire ATP-molekyler.
Se denne videoen for å lære om glykolyse:
Glykolyse kan deles inn i to faser: energikrevende (også kalt kjemisk priming) og energi gir. Den første fasen er den energikrevende fasen, så det krever to ATP-molekyler for å starte reaksjonen for hvert molekyl av glukose. Imidlertid produserer slutten av reaksjonen fire Atp, noe som resulterer i en netto gevinst på to ATP-energimolekyler.
Glykolyse kan uttrykkes som følgende ligning:
\text{Glucose}+2\text{ATP}+2\text{NAD}^{+}+4\text{ADP}+2\text{P}_{i}\to\text{Pyruvate}+4\text{ATP}+2\text{NADH}+2\text{H}^{+}p denne ligningen sier at glukose, i kombinasjon MED ATP (energikilden), NAD+ (et koenzym som fungerer som en elektron—akseptor) og uorganisk fosfat, bryter ned i to pyruvatmolekyler, og genererer fire ATP—molekyler-for et netto utbytte på TO ATP-og to energiholdige NADH-koenzymer. NADH som produseres i denne prosessen vil bli brukt senere for å produsere ATP i mitokondriene. Ved slutten av denne prosessen genererer et glukosemolekyl to pyruvatmolekyler, to HØY-ENERGI ATP-molekyler og to elektronbærende NADH-molekyler.
følgende diskusjoner av glykolyse inkluderer enzymer som er ansvarlige for reaksjonene. Når glukose kommer inn i en celle, legger enzymet heksokinase (eller glukokinase i leveren) raskt et fosfat for å konvertere det til glukose-6-fosfat. En kinase er en type enzym som legger til et fosfatmolekyl til et substrat(i dette tilfellet glukose, men det kan også være sant for andre molekyler). Dette konverteringstrinnet krever EN ATP og fanger i hovedsak glukosen i cellen, og forhindrer den i å passere tilbake gjennom plasmamembranen, slik at glykolyse kan fortsette. Det fungerer også for å opprettholde en konsentrasjonsgradient med høyere glukose nivåer i blodet enn i vevet. Ved å etablere denne konsentrasjonsgradienten vil glukosen i blodet kunne strømme fra et område med høy konsentrasjon (blodet) til et område med lav konsentrasjon (vevet) som enten skal brukes eller lagres. Hexokinase finnes i nesten alle vev i kroppen. Glukokinase, derimot, uttrykkes i vev som er aktive når blodsukkernivået er høyt, for eksempel leveren. Heksokinase har en høyere affinitet for glukose enn glukokinase og kan derfor omdanne glukose raskere enn glukokinase. Dette er viktig når nivåene av glukose er svært lave i kroppen, da det tillater glukose å reise fortrinnsvis til de vevene som krever det mer.
i neste trinn i den første fasen av glykolyse omdanner enzymet glukose-6-fosfatisomerase glukose-6-fosfat til fruktose-6-fosfat. Som glukose er fruktose også et seks karbonholdig sukker. Enzymet phosphofructokinase – 1 legger deretter til en mer fosfat for å konvertere fruktose-6-fosfat til fruktose-1-6-bisfosfat, et annet seks-karbon sukker, ved hjelp AV et ANNET ATP-molekyl. Aldolase bryter deretter ned denne fruktosen-1-6-bisfosfat i to tre-karbon molekyler, glyceraldehyd-3-fosfat og dihydroksyaceton fosfat. Triosefosfatisomeraseenzymet omdanner deretter dihydroksyacetonfosfat til et andre glyceraldehyd-3-fosfatmolekyl. Derfor, ved slutten av denne kjemisk – priming eller energikrevende fase, brytes et glukosemolekyl ned i to glyceraldehyd-3-fosfatmolekyler.
den andre fasen av glykolyse, den energiavkastende fasen, skaper energien som er produktet av glykolyse. Glyceraldehyd-3-fosfat dehydrogenase omdanner hver tre-karbon glyceraldehyd-3-fosfat produsert under
energikrevende fase til 1,3-bisfosfoglyserat. Denne reaksjonen frigjør et elektron som deretter hentes AV NAD+ for å skape ET NADH-molekyl. NADH er et høyenergimolekyl, som ATP, men i motsetning TIL ATP, brukes det ikke som energivaluta av cellen. Fordi det er to glyceraldehyd-3-fosfatmolekyler, syntetiseres to NADH-molekyler under dette trinnet. Hvert 1,3-bisfosfoglyserat blir deretter defosforylert (dvs. et fosfat fjernes) av fosfoglyseratkinase til 3-fosfoglyserat. Hvert fosfat frigjort i denne reaksjonen kan konvertere ETT molekyl ADP til et HØY-ENERGI ATP-molekyl, noe som resulterer i en gevinst på to ATP-molekyler.
enzymet fosfoglyceratmutase omdanner deretter 3-fosfoglyceratmolekylene til 2-fosfoglycerat. Enolaseenzymet virker deretter på 2-fosfoglyseratmolekylene for å konvertere dem til fosfoenolpyruvatmolekyler. Det siste trinnet av glykolyse innebærer defosforylering av de to fosfoenolpyruvatmolekylene ved pyruvatkinase for å skape to pyruvatmolekyler og to ATP-molekyler.i sammendraget bryter ett glukosemolekyl ned i to pyruvatmolekyler, og skaper to netto ATP-molekyler og to NADH-molekyler ved glykolyse. Derfor genererer glykolyse energi for cellen og skaper pyruvatmolekyler som kan behandles videre gjennom den aerobiske Krebs syklusen (også kalt sitronsyresyklusen eller trikarboksylsyresyklusen); omdannes til melkesyre eller alkohol (i gjær) ved gjæring; eller brukes senere for syntese av glukose gjennom glukoneogenese.
Anaerob Respirasjon
når oksygen er begrenset eller fraværende, går pyruvat inn i en anaerob vei. I disse reaksjonene kan pyruvat omdannes til melkesyre. I tillegg til å generere en ekstra ATP, tjener denne banen til å holde pyruvatkonsentrasjonen lav, slik at glykolyse fortsetter, og det oksiderer NADH i nad + som trengs av glykolyse. I denne reaksjonen erstatter melkesyre oksygen som den endelige elektronacceptoren. Anaerob respirasjon forekommer i de fleste celler i kroppen når oksygen er begrenset eller mitokondrier er fraværende eller ikke-funksjonelle. For eksempel, fordi erytrocytter (røde blodlegemer) mangler mitokondrier, må de produsere ATP fra anaerob respirasjon. DETTE er en effektiv VEI FOR ATP-produksjon i korte perioder, alt fra sekunder til noen få minutter. Melkesyren som produseres diffunderer inn i plasma og føres til leveren, hvor Den omdannes tilbake til pyruvat eller glukose via Cori-syklusen. På samme måte, når en person trener, bruker muskler ATP raskere enn oksygen kan leveres til dem. De er avhengige av glykolyse og melkesyreproduksjon for rask ATP-produksjon.
Aerob Respirasjon
i nærvær av oksygen kan pyruvat komme inn I Krebs-syklusen hvor ytterligere energi ekstraheres som elektroner overføres FRA pyruvatet TIL reseptorene NAD+, BNP OG FAD, med karbondioksid som et «avfallsprodukt» (Figur 3). NADH og FADH2 sender elektroner videre til elektrontransportkjeden, som bruker den overførte energien til å produsere ATP. Som terminaltrinnet i elektrontransportkjeden er oksygen den terminale elektronacceptoren og skaper vann inne i mitokondriene.
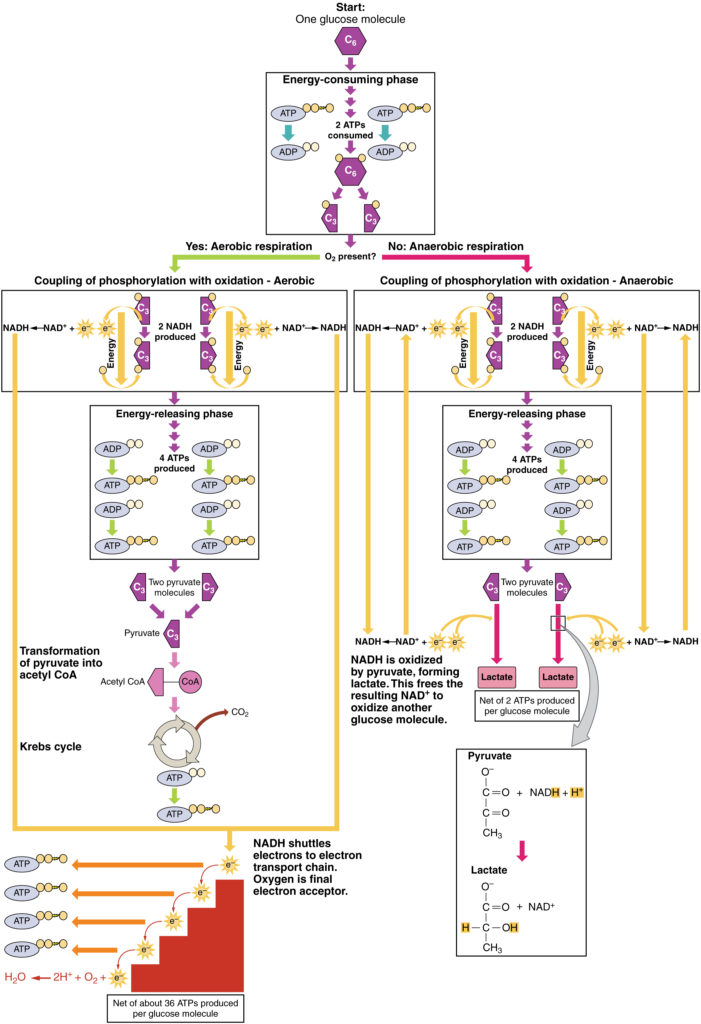
Figur 3. Klikk for å se et større bilde. Prosessen med anaerob respirasjon omdanner glukose til to laktatmolekyler i fravær av oksygen eller i erytrocytter som mangler mitokondrier. Under aerob respirasjon oksyderes glukose i to pyruvatmolekyler.
Krebs Syklus/Sitronsyre Syklus/Trikarboksylsyre Syklus
pyruvatmolekylene som genereres under glykolyse, transporteres over mitokondriamembranen til den indre mitokondriamatrisen, hvor de metaboliseres av enzymer i En vei som kalles Krebs syklusen (Figur 4). Krebs syklusen kalles også ofte sitronsyresyklusen eller tricarboxylsyre (TCA) syklusen. Under Krebs syklusen opprettes høyenergimolekyler, inkludert ATP, NADH OG FADH2. NADH og FADH2 sender deretter elektroner gjennom elektrontransportkjeden i mitokondriene for å generere FLERE ATP-molekyler.
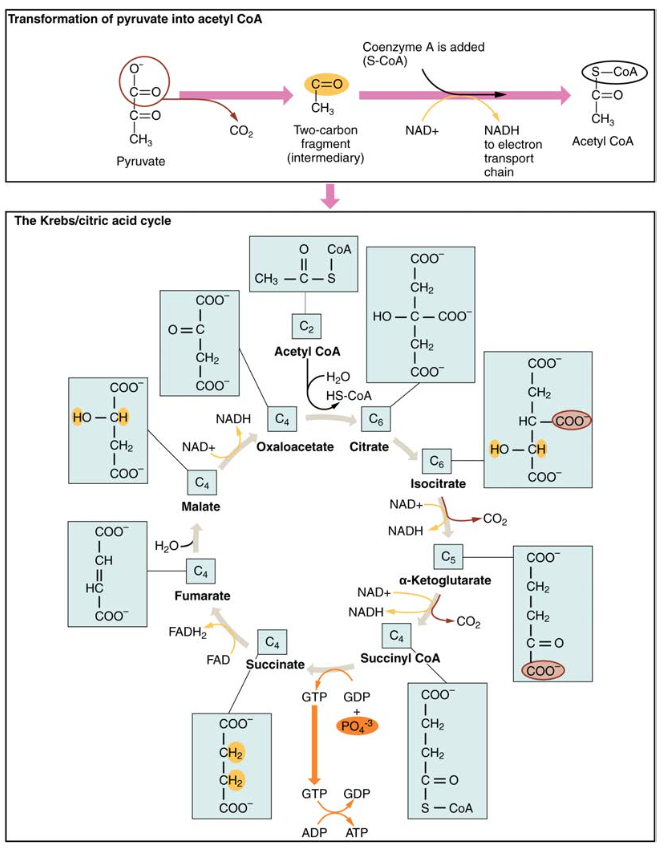
Figur 4. Under Krebs syklusen omdannes hvert pyruvat som genereres av glykolyse til et to-karbon acetyl CoA-molekyl. Acetyl CoA blir systematisk behandlet gjennom syklusen og produserer HØY-energi NADH, FADH2 og ATP molekyler.
Se denne animasjonen for å observere Krebs syklusen.det tre-karbon pyruvatmolekylet som genereres under glykolyse beveger seg fra cytoplasma til mitokondriellmatrisen, hvor det omdannes av enzymet pyruvat dehydrogenase til et to-karbon acetyl koenzym A (acetyl CoA) molekyl. Denne reaksjonen er en oksidativ dekarboksyleringsreaksjon. Den omdanner tre-karbon pyruvat til et to-karbon acetyl CoA molekyl, frigjør karbondioksid og overfører to elektroner som kombinerer MED NAD+ for å danne NADH. Acetyl CoA går Inn I Krebs-syklusen ved å kombinere med et fire-karbonmolekyl, oksaloacetat, for å danne seks-karbonmolekyletsitrat eller sitronsyre, samtidig som koenzymet frigjøres et molekyl.seks-karbon citratmolekylet omdannes systematisk til et fem-karbonmolekyl og deretter et fire-karbonmolekyl, som slutter med oksaloacetat, begynnelsen av syklusen. Underveis vil hvert citratmolekyl produsere EN ATP, EN FADH2 og tre NADH. FADH2 OG NADH vil gå inn i det oksidative fosforyleringssystemet som ligger i den indre mitokondriamembranen. I Tillegg leverer Krebs-syklusen utgangsmaterialene for å behandle og bryte ned proteiner og fett.for å starte Krebs syklusen kombinerer citratsyntase acetyl CoA og oksaloacetat for å danne et seks-karbon citratmolekyl; CoA frigjøres deretter og kan kombinere med et annet pyruvatmolekyl for å starte syklusen igjen. Aconitase-enzymet omdanner citrat til isocitrat. I to påfølgende trinn med oksidativ dekarboksylering dannes to molekyler CO2 og TO NADH-molekyler når isocitrat-dehydrogenase omdanner isocitrat til fem-karbon α-ketoglutarat, som deretter katalyseres og omdannes til fire-karbon succinyl-CoA ved α-ketoglutarat dehydrogenase. Enzymet succinyl CoA dehydrogenase omdanner deretter succinyl CoA til succinat og danner høyenergimolekylet GTP, som overfører sin energi TIL ADP for å produsere ATP. Succinat dehydrogenase omdanner deretter succinat til fumarat, og danner ET MOLEKYL AV FADH2. Fumarase omdanner deretter fumarat til malat, som malatdehydrogenase omdanner deretter tilbake til oksaloacetat mens nad+ reduseres TIL NADH. Oksaloacetat er så klar til å kombinere med neste acetyl CoA for å starte Krebs syklusen igjen (Se Figur 4). For hver sving av syklusen opprettes tre NADH, EN ATP (GJENNOM GTP) og en FADH2. Hvert karbon av pyruvat omdannes TIL CO2, som frigjøres som et biprodukt av oksidativ (aerob) respirasjon.
Oksidativ Fosforylering og Elektrontransportkjeden
elektrontransportkjeden (ETC) bruker NADH og FADH2 produsert Av Krebs syklusen for å generere ATP. Elektroner FRA NADH OG FADH2 overføres gjennom proteinkomplekser innebygd i den indre mitokondriamembranen ved en rekke enzymatiske reaksjoner. Elektrontransportkjeden består av en serie av fire enzymkomplekser (Kompleks I-Kompleks IV) og to koenzymer (ubiquinon og Cytokrom c), som fungerer som elektronbærere og protonpumper som brukes Til å overføre H+ – ioner inn i rommet mellom indre OG ytre mitokondriemembraner (Figur 5). ETC par overføring av elektroner mellom en donor (SOM NADH) og en elektron akseptor (Som O2) med overføring av protoner (H+ ioner) over den indre mitokondriamembranen, slik at prosessen med oksidativ fosforylering. I nærvær av oksygen sendes energi trinnvis gjennom elektronbærerne for gradvis å samle den energien som trengs for å feste et fosfat TIL ADP og produsere ATP. Rollen av molekylært oksygen, O2, er som terminal elektron akseptor FOR ETC. Dette betyr at når elektronene har passert gjennom hele ETC, må de sendes til et annet, separat molekyl. Disse elektronene, o2-og H+ – ionene fra matrisen kombineres for å danne nye vannmolekyler. Dette er grunnlaget for ditt behov for å puste inn oksygen. Uten oksygen opphører elektronstrømmen GJENNOM ETC.
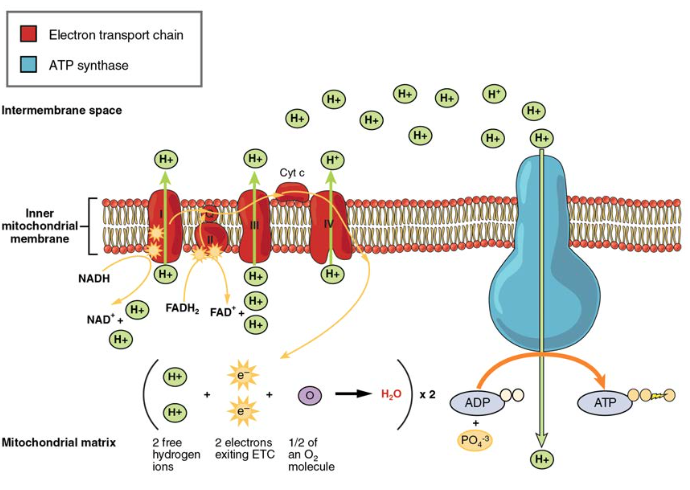
figur 5. Elektrontransportkjeden er en serie elektronbærere og ionpumper som brukes Til å pumpe H + – ioner ut av den indre mitokondrielle matrisen.
Se denne videoen for å lære om elektrontransportkjeden.
elektronene frigjort FRA NADH OG FADH2 føres langs kjeden av hver av bærerne, som reduseres når de mottar elektronen og oksyderes når de sender den videre til neste bærer. Hver av disse reaksjonene frigjør en liten mengde
energi, som brukes Til å pumpe H + – ioner over den indre membranen. Akkumuleringen av disse protonene i rommet mellom membranene skaper en protongradient med hensyn til mitokondriell matrise.også innebygd i den indre mitokondriamembranen er et fantastisk proteinporekompleks kalt ATP-syntase. Effektivt er det en turbin som drives av strømmen Av H + – ioner over den indre membranen ned en gradient og inn i mitokondriell matrise. Når H + – ionene krysser komplekset, roterer akselen til komplekset. Denne rotasjonen gjør det mulig for ANDRE DELER AV ATP-syntase å oppmuntre ADP og Pi til å skape ATP. Ved regnskapsføring av totalt ANTALL ATP produsert per glukosemolekyl gjennom aerob respirasjon, er det viktig å huske følgende punkter:
- et nett av to ATP produseres gjennom glykolyse (fire produsert og to forbrukes i energikrevende stadium). IMIDLERTID brukes disse TO ATP til å transportere NADH produsert under glykolyse fra cytoplasma til mitokondriene. Derfor er nettoproduksjonen AV ATP under glykolyse null.I alle faser etter glykolyse må ANTALLET ATP, NADH OG FADH2 som produseres multipliseres med to for å reflektere hvordan hvert glukosemolekyl produserer to pyruvatmolekyler.
- I ETC produseres omtrent tre ATP for hver oksidert NADH. IMIDLERTID produseres bare om lag to ATP for hver oksidert FADH2. Elektronene fra FADH2 produserer mindre ATP, fordi de starter på et lavere punkt I ETC (Kompleks II) sammenlignet med elektronene FRA NADH (Kompleks I) (Figur 5).
for hvert glukosemolekyl som går inn i aerob respirasjon, produseres derfor en netto total på 36 ATPs (Se Figur 6).
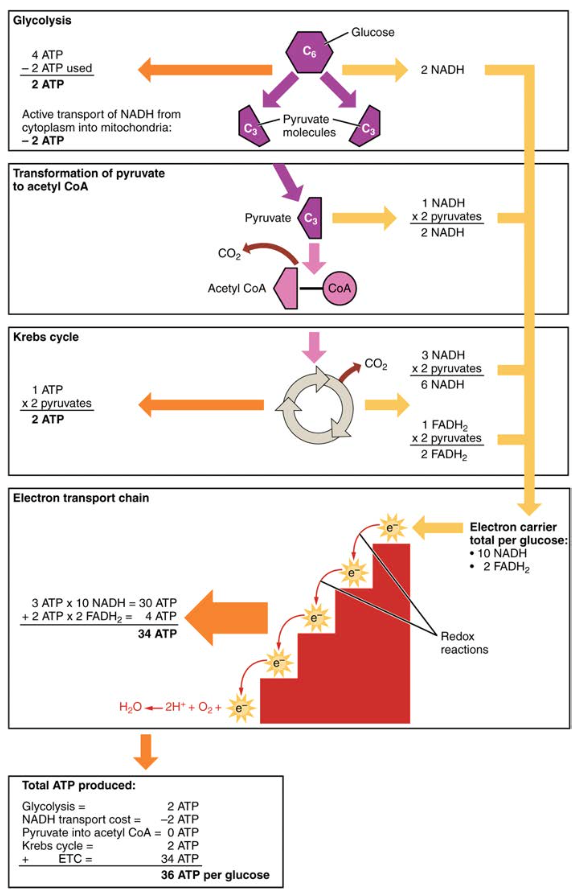
Figur 6. Karbohydratmetabolismen innebærer glykolyse, Krebs-syklusen og elektrontransportkjeden.
Glukoneogenese
Glukoneogenese er syntesen av nye glukosemolekyler fra pyruvat, laktat, glyserol eller aminosyrene alanin eller glutamin. Denne prosessen foregår hovedsakelig i leveren i perioder med lav glukose, det vil si under faste, sult og lavt karbohydrat dietter. Så, spørsmålet kan heves på hvorfor kroppen ville skape noe den nettopp har brukt en god del innsats for å bryte ned? Visse viktige organer, inkludert hjernen, kan bare bruke glukose som energikilde; derfor er det viktig at kroppen opprettholder en minimum blodglukosekonsentrasjon. Når blodglukosekonsentrasjonen faller under det bestemte punktet, syntetiseres ny glukose i leveren for å øke blodkonsentrasjonen til normal.
Glukoneogenese er ikke bare det motsatte av glykolyse. Det er noen viktige forskjeller (Figur 7). Pyruvat er et vanlig utgangsmateriale for glukoneogenese. For det første omdannes pyruvatet til oksaloacetat. Oksaloacetat tjener deretter som substrat for enzymet fosfoenolpyruvatkarboksykinase (PEPCK), som omdanner oksaloacetat til fosfoenolpyruvat (PEP). Fra dette trinnet er glukoneogenese nesten motsatt av glykolyse. PEP omdannes tilbake til 2-fosfoglyserat, som omdannes til 3-fosfoglyserat. Deretter omdannes 3-fosfoglyserat til 1,3 bisfosfoglyserat og deretter til glyceraldehyd-3-fosfat. To molekyler glyceraldehyd-3-fosfat kombineres deretter for å danne fruktose-1-6-bisfosfat, som omdannes til fruktose 6-fosfat og deretter til glukose-6-fosfat. Til slutt genererer en rekke reaksjoner glukose selv. I glukoneogenese (sammenlignet med glykolyse) erstattes enzymet heksokinase med glukose-6-fosfatase, og enzymet fosfofructokinase – 1 erstattes av fruktose-1,6-bisfosfatase. Dette hjelper cellen til å regulere glykolyse og glukoneogenese uavhengig av hverandre.som det vil bli diskutert som en del av lipolyse, kan fett brytes ned i glyserol, som kan fosforyleres for å danne dihydroksyacetonfosfat eller DHAP. DHAP kan enten gå inn i glykolytisk vei eller brukes av leveren som substrat for glukoneogenese.
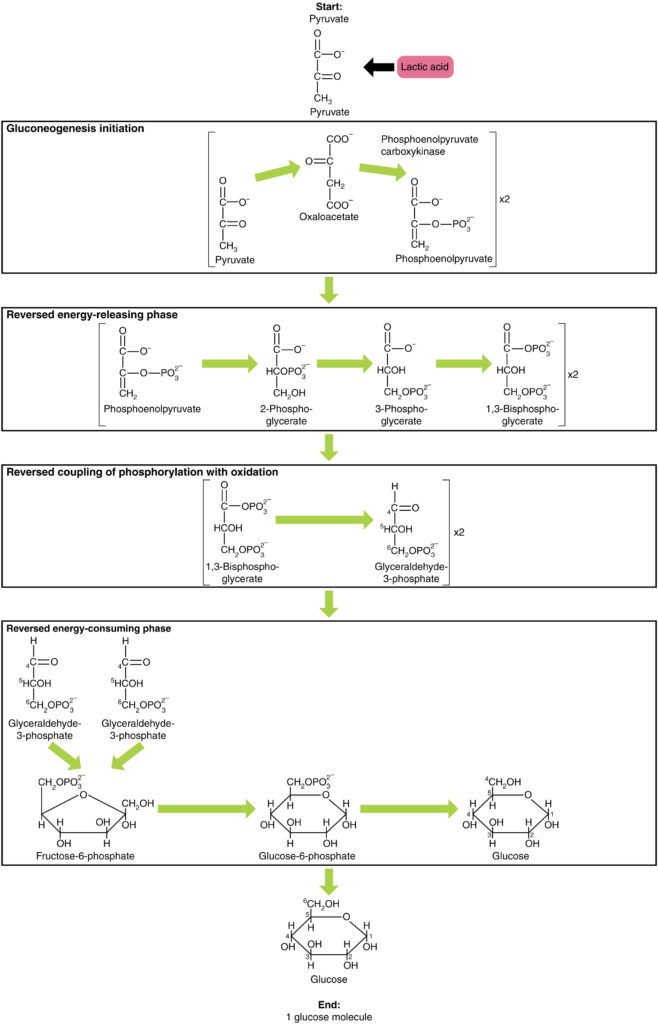
Figur 7. Klikk for å se et større bilde. Glukoneogenese er syntesen av glukose fra pyruvat, laktat, glyserol, alanin eller glutamat.
Aldring og Kroppens Metabolic Rate
den menneskelige kroppens metabolic rate avtar nesten 2 prosent per tiår etter alder 30. Endringer i kroppssammensetning, inkludert redusert muskelmasse, er for det meste ansvarlig for denne reduksjonen. Den mest dramatiske tap av muskelmasse, og påfølgende nedgang i metabolic rate, oppstår mellom 50 og 70 år. Tap av muskelmasse er ekvivalent med redusert styrke, som har en tendens til å hemme eldre fra å engasjere seg i tilstrekkelig fysisk aktivitet. Dette resulterer i en positiv-feedback system der redusert fysisk aktivitet fører til enda mer muskel tap, ytterligere redusere metabolismen.Det er flere ting som kan gjøres for å forhindre generell nedgang i metabolisme og å kjempe tilbake mot den sykliske naturen av disse nedgangen. Disse inkluderer å spise frokost, spise små måltider ofte, spise mye magert protein, drikkevann for å forbli hydrert, trene (inkludert styrketrening) og få nok søvn. Disse tiltakene kan bidra til å holde energinivået fra å slippe og dempe trangen til økt kaloriforbruk fra overdreven snacking. Selv om disse strategiene ikke er garantert å opprettholde metabolisme, bidrar de til å forhindre muskel tap og kan øke energinivået. Noen eksperter foreslår også å unngå sukker, noe som kan føre til overflødig fettlagring. Krydret mat og grønn te kan også være gunstig. Fordi stress aktiverer kortisolfrigivelse, og kortisol bremser stoffskiftet, kan det også hjelpe å unngå stress, eller i det minste praktisere avslappeteknikker.
Kapittel Gjennomgang
Metabolske enzymer katalyserer katabolske reaksjoner som bryter ned karbohydrater i mat. Energien som frigjøres brukes til å drive cellene og systemene som utgjør kroppen din. Overflødig eller unutilized energi lagres som fett eller glykogen for senere bruk. Karbohydratmetabolismen begynner i munnen, hvor enzymet spyttamylase begynner å bryte ned komplekse sukkerarter i monosakkarider. Disse kan deretter transporteres over tarmmembranen i blodet og deretter til kroppsvev. I cellene behandles glukose, et seks-karbon sukker, gjennom en sekvens av reaksjoner i mindre sukkerarter, og energien lagret inne i molekylet frigjøres. Det første trinnet med karbohydratkatabolisme er glykolyse, som produserer pyruvat, NADH og ATP. Under anaerobe forhold kan pyruvatet omdannes til laktat for å holde glykolyse i arbeid. Under aerobe forhold går pyruvat Inn I Krebs syklusen, også kalt sitronsyresyklusen eller tricarboxylsyre syklusen. I TILLEGG til ATP produserer Krebs-syklusen HØY-energi FADH2-og NADH-molekyler, som gir elektroner til den oksidative fosforyleringsprosessen som genererer MER HØY-ENERGI ATP-molekyler. For hvert molekyl av glukose som behandles i glykolyse, kan et nett på 36 ATPs opprettes ved aerob respirasjon.
UNDER anaerobe forhold er ATP-produksjonen begrenset til de som genereres av glykolyse. Mens totalt fire Atp-er produseres av glykolyse, er det nødvendig med to for å starte glykolyse, så det er et nettoutbytte av to ATP-molekyler.under forhold med lav glukose, som fasting, sult eller lavt karbohydrat dietter, kan glukose syntetiseres fra laktat, pyruvat, glyserol, alanin eller glutamat. Denne prosessen, kalt glukoneogenese, er nesten omvendt av glykolyse og tjener til å skape glukosemolekyler for glukoseavhengige organer, som hjernen, når glukosenivået faller under normalt.
Selvkontroll
Svar på spørsmålet (e) nedenfor for å se hvor godt du forstår emnene dekket i forrige avsnitt.
Kritisk Tenkning Spørsmål
- Forklar hvordan glukose metaboliseres for Å gi ATP.
- Diskuter mekanismen cellene bruker for å skape en konsentrasjonsgradient for å sikre kontinuerlig opptak av glukose fra blodet.
Ordliste
polysakkarider: komplekse karbohydrater som består av mange monosakkarider
monosakkarid: minste, monomere sukkermolekyl
spyttamylase: cellulær respirasjon: PRODUKSJON AV ATP fra glukoseoksidasjon via glykolyse, Krebs-syklusen og oksidativ fosforylering
glykolyse: serie metabolske reaksjoner som bryter ned glukose i pyruvat og produserer ATP
pyruvat: tre-karbon sluttprodukt av glykolyse og utgangsmateriale som omdannes til acetyl CoA som går inn i
Krebs syklus: også kalt sitronsyresyklusen eller tricarboxylsyre syklusen, omdanner pyruvat TIL CO2 og HØY-energi FADH2, NADH OG ATP molekyler
sitronsyre syklus eller tricarboxylsyre syklus (TCA): også kalt Krebs syklusen eller tricarboxylsyre syklusen; omdanner pyruvat TIL CO2 og HØY-energi FADH2, NADH OG ATP molekyler
energikrevende fase, første fase av glykolyse, hvor TO molekyler AV ATP er nødvendig å starte reaksjonen
glukose-6-fosfat: fosforylert GLUKOSE PRODUSERT I FØRSTE trinn AV GLYKOLYSE
heksokinase: cellulært enzym, funnet i de fleste vev, som omdanner glukose til glukose-6-fosfat ved opptak i cellen
Glukokinase: cellularenzyme, funnet i leveren, som omdanner glukose til glukose-6-fosfat ved opptak i cellen
energigivende fase: andre fase av glykolyse, hvor energi produseres
terminal elektron akseptor: ATP produksjonsvei hvor elektroner føres gjennom en rekke oksidasjonsreduksjonsreaksjoner som danner vann og produserer en protongradient
krebs cycle: også kalt sitronsyresyklusen eller trikarboksylsyresyklusen, konverterer pyruvat TIL CO2 og HØY-energi FADH2, NADH OG ATP molekyler
elektrontransportkjede (ETC): ATP-produksjonsvei hvor elektroner føres gjennom en rekke oksidasjonsreduksjonsreaksjoner som danner vann og produserer en protongradient
oksidativ fosforylering: prosess som omdanner HØY-energi NADH og FADH2 TIL ATP
ATP syntase protein: pore kompleks som skaper ATP
/p>
glukoneogenese: prosess med glukosesyntese FRA pyruvat ELLER ANDRE MOLEKYLER