Introduksjon
Fossile ressurser er det viktigste råstoffet for energi og organiske forbindelser, og deres bruk resulterer i utslipp av klimagasser knyttet til klimaendringer. Den kommende klimakrisen krever en presserende overgang fra en fossilbasert til en biobasert økonomi der lignocellulosisk biomasse i stedet for olje brukes til produksjon av drivstoff, kjemikalier og materialer. Tre er en viktig kilde til lignocellulosisk biomasse; den består hovedsakelig av sekundære fortykkede cellevegger rik på cellulose, hemicelluloser og lignin. Alle tre polymerer kan bli valorized i biobasert økonomi. Cellulose er en kilde for masse-og papirindustrien, og både cellulose og hemicelluloser kan depolymeriseres til deres monosakkarider for gjæring i f.eks. bioetanol, melkesyre og vaskemidler (Vanholme et al.( 2013b). Som lignin negativt påvirker effektiviteten av treforedling mot disse programmene, kan trær være konstruert for å akkumulere mindre lignin, for å bli mer mottagelig for produksjon av papir og gjærbare sukker. På den annen side blir lignin i økende grad ansett som en verdifull komponent i den biobaserte økonomien. Faktisk, gitt at lignin er Den største fornybare aromatiske kilden på Jorden, kan den økonomiske levedyktigheten til et bioraffineri økes betydelig hvis lignin også er verdsatt og brukt som ressurs for produksjon av kjemikalier (Holladay et al., 2007; Tuck et al., 2012; Davis et al., 2013; Ragnhild et al., 2014; Li C. et al., 2015; Van Den Bosch et al., 2015; Rinaldi et al., 2016; Upton og Kasko, 2016; Schutyser et al., 2018).ligninpolymeren består av monolignoler som produseres av fenylpropanoid-og monolignolbiosyntetiske veier, ved en rekke enzymatiske reaksjoner som begynner med deaminering av fenylalanin (Figur 1). Monolignolene syntetiseres i cytoplasma og translokeres til apoplasten, hvor de dehydrogeneres til monolignolradikaler ved hjelp av laccases og peroxidaser (Berthet et al., 2011; Zhao et al., 2013). Disse monolignolradikalene parer seg deretter med hverandre på en kombinatorisk måte, og genererer en rekke kjemiske bindinger som aryl-eterbindingen (β-O-4), resinolbindingen (β-β) og fenylkoumaranbindingen (β-5) (Boerjan et al., 2003; Ralph et al., 2004; Vanholme et al., 2010). De vanligste monolignolene er hydroksykinnamylalkoholene p-kumaryl, koniferyl og sinapylalkoholer, som genererer h -, G-og s-enhetene ved deres innlemmelse i henholdsvis ligninpolymeren (Bonawitz og Chapple, 2010; Ralph et al., 2019; Vanholme et al., 2019). Det relative bidraget av lignin byggesteinene varierer mellom taxa, utviklingsstadiet, vev og celletype, og selv cellevegglaget; lignin fra bartre (gymnosperms) består nesten utelukkende Av g enheter Med en mindre brøkdel Av H enheter, mens lignin fra hardtre (angiosperms) har s enheter i Tillegg Til G enheter og spor Av H enheter (Boerjan et al., 2003; Vanholme et al., 2010, 2019). I tillegg til disse tradisjonelle monolignolene kan en rekke andre p-hydroksylerte aromatiske molekyler inkorporeres i ligninpolymeren til forskjellige nivåer(Vanholme et al., 2019).
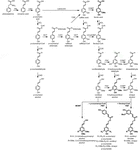
Figure 1. Lignin biosynthetic pathway. Alternative monomers and heterologously expressed enzymes are shown in bold. PAL, phenylalanine ammonia-lyase; C4H, cinnamate 4-hydroxylase; 4CL, 4-coumarate:CoA ligase; HCT, p-Hydroxycinnamoyl-CoA:quinate/shikimate-p-hydroxycinnamoyltransferase; C3’H, p-coumaroyl quinate/shikimate 3’-hydroxylase; CSE, caffeoyl shikimate esterase; CCoAOMT, caffeoyl-CoA O-methyltransferase; CCR, cinnamoyl-CoA reductase; F5H, ferulate 5-hydroxylase/CAld5H, coniferaldehyde 5-hydroxylase; COMT, koffeinsyre O-metyltransferase; CAD, cinnamylalkohol dehydrogenase; fmt, feruloyl-CoA monolignol transferase; PMT, p-kumaroyl-CoA monolignol transferase.Gitt at lignin er en viktig recalcitrance faktor i tre delignification prosesser, har store forskningsinnsats vært viet til å avdekke lignin biosyntetiske veien, og for å studere effekten av perturbations av lignin biosyntese gener på lignin mengde og sammensetning, og på tre behandling effektivitet. Mens modifikasjoner i gener som spenner fra de som koder for transkripsjonsfaktorer opp til de som koder for oksidative enzymer, har resultert i endret lignininnhold, sammensetning eller avsetning (Eriksson et al., 2000; Li Y. H. et al., 2003; Liang et al., 2008; Lu et al., 2013; Lin Et al., 2016; Xu et al., 2017; Jan et al., 2017; Obudulu et al., 2018), vil denne vurderingen fokusere på resultatene oppnådd ved å konstruere ligninbiosyntetiske gener.
Prosjektering Av Ligninveien
Tabell 1 gir en oversikt over de ulike studiene på nedregulerte eller muterte ligninbiosyntetiske gener i poppel, furu, eukalyptus og bjørk, med de resulterende effektene på trebearbeidingseffektivitet, når det bestemmes. Reduksjon av aktiviteten i et hvilket som helst trinn av ligninbiosyntetisk vei, fra PAL opp til CAD, kan resultere i reduksjon av lignininnholdet (Tabell 1). Flere parametere påvirker graden av ligninreduksjon, for eksempel målgenet og graden av nedregulering av enzymaktiviteten, som igjen avhenger av effektiviteten av silencingskonstruksjonen som brukes, størrelsen på genfamilien og redundans i genfamilien. Generelt, nedregulering av trinnene FRA C4H OPP TIL CCR resulterer i en mer dramatisk reduksjon i lignin mengde (Hu et al., 1999; Meyermans et al., 2000; Zhong et al., 2000; Li L. et al., 2003; Jia et al., 2004; Lu et al., 2004; Et al., 2007; Coleman et al., 2008a, b; Bjurhager et al., 2010; Mansfield et al., 2012; Ralph et al., 2012; Min et al., 2014; Van Acker et al., 2014; Zhou et al., 2015, 2018; Saleme et al., 2017; Jørgen et al., 2017) enn nedregulering AV F5H, COMT OG CAD (Van Doorsselaere et al., 1995; Baucher et al., 1996; Lapierre et al., 1999; Jouanin et al., 2000; Van Acker et al., 2014; Wang et al., 2018). Lignin reduksjon kan være forbundet med en økning I S / G, slik Som I C3 ‘ H- (Coleman et al., 2008a; Ralph et al., 2012) Og CCoAOMT-nedregulerte trær (Meyermans et al., 2000), eller en reduksjon I S / G-forhold som I CSE – (Saleme et al., 2017), OG COMT-downregulated trær (Van Doorsselaere et al., 1995; Lapierre et al., 1999; Jouanin et al., 2000). Interessant, lav-lignin 4CL-downregulated poplars ble funnet å ha en økning I S / G(Min et al., 2014; Xiang et al., 2017), en nedgang I S / G (Voelker et al., 2010; Zhou et al., 2015), eller forhold som kan sammenlignes med villtype (Hu et al., 1999; Li L. et al., 2003). Denne variansen kan ikke knyttes til promotoren eller metoden som brukes for nedregulering, noe som tyder på at forskjeller i graden av silencing, vekstforhold eller utviklingsstatus påvirker dette trekket. På den annen side har de sterkeste effektene På h/G / S ligninsammensetning blitt observert for trær nedregulert I C3 ‘ H og HCT, som avsetter lignin med store økninger I h-enhetsinnhold (Coleman et al., 2008a; Ralph et al., 2012; Vanholme et al., 2013a), mens trær som overexpress F5H produsere lignin sterkt beriket I s enheter (Franke et al., 2000; Li L. et al., 2003; Stewart et al., 2009), og tr r som er downregulated I COMT har dramatisk redusert s enhet biosyntese (Van Doorsselaere et al., 1995; Lapierre et al., 1999).

Tabell 1. Oversikt over skogstrær med modifisert uttrykk for ligninbiosyntesegener.
prosesseringseffektiviteten til biomassen kan også modifiseres ved økt inkorporering av molekyler som generelt representerer mindre komponenter i lignin av villtype planter. Inkorporeringen av ferulsyre i CCR-mangelfulle trær resulterer i dannelse av acetalbindinger i ligninpolymeren, som lett spaltes i sure biomasseforbehandlinger (Leplé et al., 2007; Ralph et al., 2008; Van Acker et al., 2014). Faktisk korrelerte nivåene av ferulsyre i lignin positivt med en høyere sakkarifiseringseffektivitet (Van Acker et al., 2014). Inkorporering av 5-hydroksykoniferylalkohol og 5-hydroksykoniferaldehyd i lignin AV COMT-mangelfulle poplars (Van Doorsselaere et al., 1995; Lapierre et al., 1999; Jouanin et al., 2000; Morreel et al., 2004; Lu et al., 2010) gir opphav til benzodioksanbindinger, som potensielt forhindrer kovalente bindinger mellom lignin og polysakkaridhydroksylgruppene (Weng et al., 2010; Vanholme et al., 2012; Nishimura et al., 2018). PÅ DEN annen side resulterer comt-mangel også i et mer kondensert lignin på grunn av relativt høyere nivåer av kondenserte β-β og β-5-obligasjoner, og de lavere nivåene av β-O-4-obligasjoner når s-frekvensen faller. Kjemisk pulping av tre avledet fra poplars sterkt nedregulert FOR COMT resulterte i et høyere masseutbytte, motvektet av rest lignininnholdet i massen. Disse trærne hadde et lavere lignin og et høyere celluloseinnhold (Jouanin et al., 2000). På den annen side hadde popler som var beskjedent nedregulert FOR COMT en stor nedgang i masseutbyttet, antagelig fordi lignininnholdet hadde forblitt normalt mens ligninet hadde en høyere frekvens av kondenserte bindinger som negativt påvirket ligninekstraksjonen (Lapierre et al ., 1999; Pilatus et al., 2002). Inkorporeringen av cinnamaldehyder i ligninpolymeren i CAD-mangelfulle trær resulterer i kortere ligninpolymerkjeder, og dermed en høyere andel av frie fenoliske endegrupper som øker oppløseligheten av polymeren i alkali. Inkorporeringen av cinnamaldehyder i ligninpolymeren reduserer antagelig også den kovalente interaksjonen av den alifatiske kjeden med hemicellulose, noe som igjen gjør ligninet mer løselig. I tillegg, på grunn av det utvidede konjugerte systemet som genereres når en cinnamaldehyd β-o-4 par med en annen monomer, blir den aromatiske eterbindingen av det innlemmede cinnamaldehyd mer utsatt for alkalisk spaltning (Lapierre et al., 1989; Van Acker et al., 2017).Ligninpolymerisering er en kombinatorisk radikal koblingsprosess, slik at et bredt spekter av fenolforbindelser naturlig kan inkorporeres i ligninpolymeren (Boerjan et al., 2003; Vanholme et al., 2019). Forskere har forsøkt å skreddersy ligninmengden og sammensetningen for å forbedre biomassebehandling ved uttrykk for heterologe gener, med sikte på biosyntese og inkorporering av forskjellige kompatible fenolforbindelser som alternative monolignoler i ligninpolymeren (Ralph, 2006; Vanholme et al., 2012; Mottiar et al.(2016; Mahon Og Mansfield, 2018). Et eksempel er innføring av gener som koder enzymer som er nødvendig For s enhet biosyntese i furu; samtidig ekspresjon AV F5H, COMT OG CAD vellykket innført s enheter I Pinus radiata (Wagner et al., 2015; Edmunds et al., 2017). Innføringen av genet som koder for en monolignol 4-o-metyltransferase (MOMT4) i poppel fører til dannelsen av 4-o-metylerte nåletrær og sinapylalkoholer, som ikke kan inkorporeres i den voksende ligninpolymeren fordi de mangler den aromatiske hydroksylgruppen. Dette fører til en stopp i lignin polymerisering og resulterer i trær med lavere lignin innhold og høyere sakkarifisering effektivitet (Bhuiya Og Liu, 2010; Cai et al ., 2016). Poplars har også blitt konstruert for å inneholde esterbindinger i ligninpolymerens ryggrad. Coniferyl ferulate estere ble innført i polymeren via ekspresjon AV EN FERULOYL-CoA: MONOLIGNOL TRANSFERASE (FMT) gen avledet Fra Angelica sinensis (Wilkerson et al., 2014), som fører til en forbedret sakkarifiseringseffektivitet under ulike forbehandlingsforhold (Wilkerson et al., 2014; Kim et al., 2017; Jørgen et al., 2018), og en forbedret kraftpulpeffektivitet sammenlignet med villtype (Zhou et al., 2017). Monolignol p-kumaratestere har også blitt konstruert i poppel, via uttrykk av en ris p-KUMAROYL-CoA: MONOLIGNOL TRANSFERASE (PMT) gen, noe som resulterer i en høyere frekvens av resistente interunit obligasjoner og en høyere frekvens Av G og s terminalenheter med frie fenoliske grupper (Smith et al., 2015; Simonsen et al., 2016). Mens I Arabidopsis resulterte det heterologe uttrykket AV PMT i en redusert ligninmengde ledsaget av økt sakkarifiseringseffektivitet (Sibout et al., 2016), var det ingen reduksjon i ligninmengde i poppel og sakkarifiseringseffektiviteten ble ikke bestemt (Smith et al., 2015).
mens flere modifikasjoner av ligninmengden og sammensetningen ble vist å gi forbedringer i biomassebehandling, ble disse modifikasjonene ofte ledsaget av en biomasseutbyttestraff (Leplé et al., 2007; Wadenbä et al., 2008; Wagner et al., 2009; Voelker et al., 2010; Stout et al., 2014; Van Acker et al., 2014; Sykes et al., 2015; Zhou et al., 2018). En nylig metastudi forstyrret 21 ligninbiosyntesegener I P. trichocarpa, og omfattende integrert resultatene av transkriptomic, proteomic, fluxomic, og phenomic data av 221 linjer. Forfatterne konkluderte med at trevekst ikke er forbundet med ligninmengde, underenhetssammensetning eller spesifikke bindinger (Wang et al., 2018), men heller korrelert med tilstedeværelsen av kollapsede xylem-fartøy (Coleman et al., 2008a, b; Wagner et al., 2009; Voelker et al., 2010; Vargas et al., 2016; De Meester et al., 2018), aktivering av en celleveggintegritetsvei (Bonawitz et al., 2014) og / eller akkumulering av kjemiske hemmere (Gallego-Giraldo et al., 2011; Muro-Villanueva et al., 2019).mens det er gjort betydelige anstrengelser for å redusere lignininnholdet ved nedregulering av ligninbiosyntetiske gener, har studier på oppregulering av ligninveien og overproduksjon av lignin vært knappe. Faktisk viser rapporter OM overuttrykk AV F5H en uendret eller til og med en reduksjon i lignininnhold (Huntley et al., 2003; Li L. et al., 2003; Stewart et al., 2009; Mansfield et al., 2012; Edmunds et al., 2017). Overekspresjonen AV CAD og COMT har resultert i gendeaktivering i stedet for oppregulering, eller ingen effekt på uttrykksnivåer ble påvist (Baucher et al., 1996; Lapierre et al., 1999; Jouanin et al., 2000; Leplé, 2007; Van Acker et al., 2014). Overuttrykket AV r2r3-MYB transkripsjonsfaktorene PtoMYB92, PtoMYB216 og PtoMYB74 resulterte alle i ekstra xylemlag, tykkere xylemcellevegger samt ektopisk ligninavsetning, og plantene akkumulerte 13-50% mer lignin (Tian Q. et al., 2013; Li C. F. et al., 2015; Li et al., 2018). MYB-overekspresjonslinjene oppregulerte konstitutivt ligninbiosynteseveien, og mens planter overuttrykker MYB92 og MYB74 hadde en biomassestraff, resulterte OVEREKSPRESJONEN AV MYB216 i planter med opptil 50% mer lignin og ingen utviklingsmessig fenotype. Som lignin blir i økende grad ansett som en viktig ressurs for bærekraftig produksjon av kjemikalier (Cao et al., 2018) prosjektering av planter overproducerende lignin bør utforskes videre.
Feltforsøk
eksemplene diskutert ovenfor viser tydelig at lignin engineering via ned-eller oppregulering av fenylpropanoid vei gener-eller uttrykk for heterologe gener – har potensial til å øke prosesseringseffektiviteten av lignocellulosic biomasse. På grunn av praktiske og regulatoriske årsaker rapporterer de fleste studier om data hentet fra analysen av trær dyrket i et drivhus. Imidlertid tar eksperimenter med trær som vokser i et drivhus vanligvis ikke hensyn til utviklingsprosesser som vekststans og dvalemodus. I tillegg gir drivhuseksperimenter ikke tilstrekkelig innsikt i samspillet mellom det konstruerte anlegget med miljøfaktorer som jordtype, vind og patogener. Å forstå disse interaksjonene er et viktig skritt i oversettelsen av forskningsresultater mot kommersielle applikasjoner. Faktisk, kroppen av arbeidet produsert av studier som tillatelse til å etablere feltforsøk ble gitt, fremhever viktige forskjeller i fenotype mellom drivhus-og felt dyrket trær. Tabell 1 oppsummerer rapportene om feltforsøk utført MED 4cl, CCoAOMT, CCR, COMT og CAD nedregulerte trær.Bekrefter potensialet for modifisert lignocellulosic biomasse som et substrat for applikasjoner, flere lignin-utviklet felt-rettssaken vokst trær viste forbedringer i treforedling. Poplars nedregulert For CCoAOMT vokst i 5 år i et feltforsøk I Beijing (Kina), viste økt glukose og xylosefrigivelse ved sakkarifisering (Wang et al., 2012). Poplars nedregulert FOR CCR og vokst i et feltforsøk I Frankrike, viste seg å være mer mottagelig for kjemisk kraftmasse (Leplé et al., 2007). To ytterligere feltforsøk utført I Frankrike og Belgia med CCR-downregulated poplars resulterte i opptil 160% forbedring i etanolproduksjon i SAMTIDIGE sakkarifisering og gjæring (SSF) analyser; men plantene hadde opptil 50% biomassereduksjon (Van Acker et al., 2014). Feltforsøk med CAD-downregulated poppel viste også lovende resultater. Disse trærne viste litt mindre lignin enn villtype og viste seg mer mottagelig for kraft delignification (Lapierre et al., 1999). Konsekvent viste de samme linjene som vokste i større feltforsøk i Frankrike og Storbritannia en mild nedgang i ligninmengde og en forbedring i kraftpulping ansett kommersielt relevant, siden plantene trengte 6% mindre alkali for å oppnå en delignifisering som ligner på villtype trær (Pilatus et al., 2002).motstridende rapporter om både biomasseutbytte og nedstrøms prosesseringseffektivitet tyder imidlertid på at disse parametrene er sterkt påvirket av miljøfaktorer. Mens en feltforsøk utført I Kina ved HJELP AV 4CL downregulated poplars fant at selv med en 28% reduksjon i lignininnhold sammenlignet med villtype, hadde trærne omtrent 8% økt høyde (Tian Xm et al., 2013), i samsvar med drivhusstudier (Hu et al., 1999), andre feltforsøk fant AT 4CL-downregulated poplars hadde redusert biomasse og ble noen ganger til og med dverget (Voelker et al., 2010; Stout et al., 2014; Marchin et al., 2017). Rapporter også avvike om nedstrøms prosesseringseffektivitet av tre avledet fra DISSE 4CL-downregulated felt-dyrket popler. Mens opptil 100% økning i sukkerutvinning ble funnet FOR 4CL1-downregulated trær (35S-drevet antisense 4CL konstruksjon) vokst i et fjellområde I Usa (Xiang et al., 2017), data hentet fra feltstudier utført I Oregon (Usa) fant At Pt4CL1 promoter-drevet antisense-drevet 4cl1-poplars ikke hadde noen forbedring i sakkarifiseringseffektivitet sammenlignet med villtype (Voelker et al ., 2010). På samme måte fant en langsiktig studie I Wenling (Kina)AT 4CL-downregulated poplars ikke viste en signifikant forbedring i sukkerutbyttet sammenlignet med villtype (Wang et al., 2012). I begge sistnevnte tilfeller viste trærne milde reduksjoner i ligninmengde som ikke omsatte til høyere prosesseringseffektivitet, potensielt på grunn av høyere konsentrasjon av ekstrakter som kunne forstyrre enzymatisk aktivitet (Voelker et al., 2010).Feltforsøk studier har vist at miljøfaktorer kan påvirke lignifisering og gjenopprette egenskaper til villtype nivåer i forhold til nivåene oppnådd når de samme plantene ble dyrket i drivhuset. Mens 4cl-downregulated trær hadde redusert lignin innhold når dyrket under drivhus forhold, analyse av de SAMME 4cl antisense popler, men dyrket i feltet, har ofte vist at lignin innholdet ble økt i forhold til drivhus-dyrket trær og noen ganger også restaurert til vill-type nivåer (Stout et al., 2014; Xiang et al., 2017). Tilsvarende var ligninnivåene mye mindre redusert I CCR-mangelfulle poplars når de ble dyrket i feltet i forhold til når de ble dyrket i drivhuset (Van Acker et al., 2014). I hvert fall FOR CCR-mangelfulle poplars er det mulig at det høyere lignifiseringsnivået av feltvoksne trær skyldes at treprøvene ble tatt om vinteren. Når treveksten opphører om høsten, har trærne fortsatt tid til å fullt ut lignifisere celleveggene når treet går inn i dvalen, sammenlignet med drivhusdyrkede trær som utvikler ny xylem kontinuerlig. Lignin sammensetning har også vist seg å variere mellom drivhus-og felt-dyrket lav-lignin trær. 4CL-downregulated poplars vokst i et felt I North Carolina hadde lignin med lavere S / G enn når de samme linjene ble dyrket i drivhuset (Stout et al., 2014).Til sammen viser disse resultatene at data hentet fra drivhusdyrkede trær ikke lett kan ekstrapoleres til feltdyrkede trær, noe som underbygger behovet for feltforsøkforsøk på forskjellige steder. Noen linjer presenterte en avkastningsstraff som gjorde dem mindre interessante for applikasjoner, og fremhevet behovet for en bedre forståelse av det molekylære grunnlaget for avkastningsstraffen og utviklingen av strategier for å overvinne dette problemet.Lignin har vist seg å spille en viktig rolle i patogenresistens (Miedes et al., 2014; Zhao Og Dixon, 2014), og det spiller en sentral rolle i å la anlegget transportere vann. Dette tyder på at lignin modifikasjoner kan ha en innvirkning på anlegget stress toleranse. Mens ytterligere undersøkelser er nødvendig for å fullt ut løse denne muligheten, nedregulering AV 4CL, COMT, OG CAD i poppel ikke dramatisk endre mate ytelsen til blad-feeding planteetere (Tiimonen et al., 2005; Brodeur-Campbell et al., 2006; Hjalten et al., 2013). Effekten av nedregulering AV COMT og CAD i poppel på plante-insekt interaksjoner har også blitt vurdert på feltvoksne trær, og det ble vist at ligninmodifiserte trær hadde normal forekomst av besøk og mating av insekter, samt normale responser på mikrobielle patogener (Pilatus et al., 2002; Halpin et al., 2007). Disse resultatene indikerer at trær med modifisert lignin ikke nødvendigvis lider mer enn villtype planter fra skadedyr og sykdommer. Likevel viste profilering av endosfærens bakterielle mikrobiom av tre høstet fra feltdyrket, CCR-downregulerte poplars skift i bakteriesamfunnet, antagelig på grunn av den endrede overflod av bestemte fenoliske metabolitter i xylem (Beckers et al., 2017).
med tanke på lignins rolle i xylemfunksjon og struktur, har vannforholdene til noen få lav ligninmodifiserte poplarer blitt vurdert. 4CL-downregulated poplars ble funnet å ha redusert hydraulisk ledningsevne, som potensielt forstyrrer planteveksten (Marchin et al., 2017). Hydrauliske stress eksperimenter med popler nedregulert FOR CCR, COMT eller CAD viste at disse anleggene hadde en lavere motstand mot kavitasjon, samtidig opprettholde normal xylem hydraulisk ledningsevne og vanntransport (Awad et al., 2012). Disse resultatene tyder på at veksten av lav-lignin mutanter kan påvirkes av vanntilgjengelighet. Som for enhver ny hybrid oppnådd fra klassisk avl, er felttester nødvendig for å evaluere feltytelse og stresstoleranse for ligninkonstruerte trær.
Utsiktene For Lignin Engineering I Skog Trær
ytelsen til lignin-engineered planter ser ut til å være sterkt påvirket av miljøforholdene. Det er imidlertid uklart om forskjellene observert mellom drivhusdyrket og feltdyrket trær, eller mellom trær dyrket på forskjellige feltsteder, skyldes forskjellige nivåer av genundertrykkelse eller fra samspill av det konstruerte trekket med miljøet (GxE). Faktisk er ustabil nedregulering en mangel på gendeaktiveringsteknikker som er basert På RNAi. Dette bevitnes ved å observere variasjon i den røde xylemfenotypen som observeres når bestemte ligninbiosyntesegener, SOM CAD, COMT eller CCR, nedreguleres. Den røde xylemfargen er ofte ikke ensartet i hele xylemet, men vises i flekker som reflekterer variable nivåer av gendeaktivering (Leplé et al., 2007; Voelker et al., 2010; Van Acker et al., 2014; Figur 2). I tillegg kan bruk av gendeaktiveringsmetoder potensielt føre til samtidig silencing av nært beslektede genfamiliemedlemmer – kanskje i ulike grader-clouding tolkingene og kamuflere effektene av nedregulering av individuelle gener.

Figur 2. Usammenhengende gene downregulation av RNAi. Usammenhengende rød xylemfenotype observert på stammer AV CCR-mangelfulle poplarer (til høyre) dyrket i et feltforsøk I Belgia. Den røde xylem angir områder AV CCR nedregulering. Tre fra villtype trær er hvitaktig(venstre).
disse problemene kan nå lett løses ved bruk AV CRISPR-baserte genredigeringsteknologier som muliggjør stabile tap av funksjonsmutasjoner (knock-outs)i bestemte målgener, slik at disseksjonen av funksjonen til individuelle gener i familier. For eksempel viste målrettingen av individuelle 4cl-genfamiliemedlemmer i poppel AT 4CL1 er relatert til lignifisering, MENS 4CL2 er involvert i proanthocyanidinproduksjon (Zhou et al., 2015). I TILLEGG til knock-out alleler, CRISPR-basert gen redigering tillater også å skape nye alleler som gir delvis reduksjon i enzymaktivitet. Dette åpner muligheten til å finjustere nivået av gjenværende enzymaktivitet og å omgå avkastningsstraffen som ofte observeres når ligninbeløpet faller under et terskelnivå. En annen lovende avenue for lignin engineering i skog trær gjort mulig GJENNOM CRISPR-basert genom engineering er samtidig redigering av flere gener (allel stabling) for å optimalisere biomasse behandling effektivitet, som eksemplifisert I Arabidopsis hvor stabling av transaldolase (tra) og comt mutasjoner, c4h og comt mutasjoner, eller 4cl og comt mutasjoner resulterte i additive og synergistiske forbedringer i sakkarifisering effektivitet (de Vries et al., 2018). Faktisk, et system tilnærming I P. trichocarpa spår at samtidig nedregulering AV PAL og CCoAOMT, eller PAL, C3 ‘ H og CCOAOMT vil vesentlig forbedre tre egenskaper og sukker utgivelse (Wang et al., 2018).BRUKEN AV CRISPR – basert genomredigering i treforbedring for cellulose – og papirindustrien og bioraffineriindustrien, samt for produksjon av plattformaromatics fra hydrogenolytisk nedbrytning av lignin, vil være mest verdifull når denne teknologien er strategisk kombinert med andre avlsteknikker (Figur 3). Faktisk finnes stor variasjon i lignin mengde og s/G sammensetning allerede i naturlige bestander av skog trær (Studer et al., 2011). Gitt at begge trekk påvirker glukosefrigivelsen ved sakkarifisering (Yoo et al., 2018), utnytte dette genetiske mangfoldet ved konvensjonell avl, hjulpet Av Genome Wide Association Studies (GWAS) (Porth et al., 2013; Fahrenkrog et al., 2017; Liu et al., 2018), Avl Med Sjeldne Defekte Alleler (BRDA) (Vanholme et al., 2013a) eller genomisk utvalg (Yin et al., 2010; Muchero et al., 2015; Pawar et al., 2018; Xie et al., 2018), er en verdifull strategi for å skaffe linjer som har forbedret trebearbeidingseffektiviteten. Når elitetrær er oppnådd ved disse avlsmetodene, er genteknologi og CRISPR-basert genredigering av spesifikke gener en meget lovende avenue for å ytterligere forbedre disse elite genotypene uten å bryte opp deres genetiske grunnlov og uten å gå gjennom lange avlssykluser. Gitt den forestående klimakrakket, har vi ikke mer tid å miste i å vedta disse nye avlsteknikkene i vårt løp mot den biobaserte økonomien.
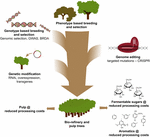
Figur 3. Genetisk forbedring av skogstrær gjennom en kombinasjon av avlsverktøy. For å akselerere den genetiske forbedringen av skogtrær for masse-og bioraffineriapplikasjoner, må klassiske og nye avlsverktøy kombineres smart. Klassisk avl innebærer fenotypisk utvalg av trær for kontrollerte kryss, etterfulgt av fenotypisk utvalg. Med fremkomsten av genomsekvensinformasjon fra mange skogtrær, har nye strategier som Genomisk Utvalg, GENOMBREDE Assosiasjonsstudier (GWAS) og Avl Med Sjeldne Defekte Alleler (BRDA) blitt utviklet for å øke hastigheten på fangst og berikelse AV DNA-polymorfier forbundet med gunstige egenskaper. CRISPR-basert genomredigering gjør det mulig å modifisere genomet på en måte som etterligner naturlige polymorfier. Genetisk modifikasjon innebærer stabil integrering av fremmed DNA i treet for å overprodusere (an) enzym (er) eller nedregulere (a) gen (er). Kombinere de klassiske Og Nye Avlsteknikker er nødvendig for å gi tilstrekkelig høy kvalitet tre for samfunnet.
Forfatterbidrag
alle forfattere oppført har gjort en betydelig, direkte og intellektuell bidrag til arbeidet, og godkjent det for publisering.
Finansiering
vi anerkjenner delvis finansiering fra IWT-SBO-prosjektet BIOLEUM (Stipend nr. 130039), OG AV SBO-FISH gjennom ARBOREF-prosjektet. AC har mottatt finansiering fra FWO og Eus Forsknings-Og Innovasjonsprogram Horizon 2020 under Marie Skł-Curie – Tilskuddsavtale nr.665501. LdV ble finansiert av Institutt for fremme Av Innovasjon Gjennom Vitenskap og Teknologi I Flandern (IWT-Vlaanderen) for en predoctoral fellowship.
Interessekonflikt
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller økonomiske forhold som kan tolkes som en potensiell interessekonflikt.
Takk
Vi takker Annick Bleys for å forberede dette manuskriptet for innsending.
Boerjan, W., Ralph, J., Og Baucher, M. (2003). Ligninbiosyntese. Annu. Rev. Plante Biol. 54, 519–546.
PubMed Abstract/Google Scholar
Lapierre, C., Pollet, B., Petit-Conil, M., Toval, G., Romero, J., Pilatus, G., Et al. (1999). Strukturelle endringer av ligniner i transgene poplars med deprimert cinnamylalkoholdehydrogenase eller koffeinsyre O-metyltransferaseaktivitet har motsatt innvirkning på effektiviteten av industriell kraftpulping. Plante Physiol. 119, 153–163.
PubMed Abstract/Google Scholar
Ralph, J. (2006). «Hva gjør en god monolignol erstatning?»I Vitenskapen og Læren Om Plantecelleveggen-Biosyntese, Struktur Og Funksjon, red. T. Hayashi (Boca Raton, FL: BrownWalker Press).
Google Scholar