Introduksjon
Vellykket samspill med vårt sensoriske miljø krever En intrikat balanse mellom to attentional seleksjonsmekanismer-den av top-down og bottom-up. Overskrift over til produsere midtgangen i ditt lokale supermarked med mål om å plukke opp noen nødvendige ingredienser for mango salat, engasjerer distribusjon av frivillig målrettet, eller top-down, attentional system slik at du aktivt søke etter alle de nødvendige ingrediensene blant de mange råvarer valg. Men, bør du høre en ringer av en mobiltelefon, det vil mest sannsynlig fange din oppmerksomhet og avbryte søket. Slike avbrudd oppstår i en bottom-up, eller stimulus-drevet, mote der bare salience av stimulus, det faktum at ringen er forskjellig fra andre lyder i miljøet, anser det verdig utvalg. Det beskrevne scenariet understreker betydningen av målrettet og stimulusdrevet utvalg for atferd, og peker på en fin balanse som må eksistere mellom de to oppmerksomhetssystemene for å forhindre «tunnelvisjon» på den ene siden og fullstendig manglende evne til å fokusere på den andre.
Top-Down Og Bottom-Up Utvalg: Behavior
Flere tiår med atferdsforskning har vært dedikert til å demonstrere at fordelingen av oppmerksomhet kan styres av observatørens intensjoner, så vel som ved salience av den fysiske stimulansen. Mye av atferdsmessige bevis for top-down og bottom-up attentional allocation har blitt gjennomgått mye andre steder (Johnston and Dark, 1986; Egeth og Yantis, 1997). For å oppsummere, viser studier som viser effekter av top-down attentional control at oppmerksomheten kan tildeles romlige steder,funksjoner, objekter, etc., etter tilstedeværelse av eksogene eller endogene signaler (Eriksen Og Hoffman, 1972; Posner, 1980; Posner et al., 1980), eller forventninger enten satt av forkunnskaper eller ved uforutsette stimulanser (Shaw, 1978; Moore Og Egeth, 1998; Geng Og Behrmann, 2002, 2005; Shomstein Og Yantis, 2004a; Drummond og Shomstein, 2010). Bevis som støtter bottom-up attentional allocation har støttet seg på ulike oppmerksomhetsfangst paradigmer, der deltakerne er engasjert i et topp-down-søk og deres oppmerksomhet omdirigeres til oppgaven-irrelevante stimuli, som viser at oppmerksomheten er fanget av funksjons enkeltfødte (unikt element; Yantis og Jonides, 1990; Theeuwes, 1991; Folk et al.( 2002) og brå utbrudd (Yantis og Jonides, 1984; Theeuwes, 1991; Koshino et al., 1992; Juola et al., 1995).Mens de fleste tidlige studier konsentrerte seg om å demonstrere bevis for top-down og bottom-up attentional selection, skiftet de siste studiene fokus til å undersøke hvordan de to attentional selection-systemene samhandler. Denne undersøkelseslinjen er drevet av observasjoner at for å effektivt velge oppgaverelevant informasjon (f. eks. ingredienser til salaten) må man aktivt hemme oppgavens irrelevante informasjon som ellers ville avlede oppmerksomheten bort fra oppgaven ved hånden. Baksiden av denne logikken er at jo mindre man er fokusert på oppgaverelatert informasjon, desto mer fangst vil følge. Det har blitt vist eksperimentelt at observatørens oppmerksomhetstilstand forutsier hvilken type informasjon, og i hvilken grad, til slutt vil fange oppmerksomhet (Folk Et al.(1992, 2002; Bacon Og Egeth, 1994; Gibson og Kelsey, 1998). For Eksempel, Folk Et al. (2002) viste at når man søker etter et rødt brev, vil en observatør lettere bli fanget av en irrelevant stimulus i periferien hvis den stimulansen er rød, eller samsvarer med målmalen på en eller annen måte. Siden observatørens topp-ned-kontrollinnstillinger er satt til å søke etter en rød funksjon, vil enhver stimulans som er rød, sannsynligvis fange oppmerksomhet og potensielt forstyrre topp-ned-kontroll. Således, med en fangstoppgave, kan oppmerksomhetssøkestrategier skille seg fra hverandre ved å variere likhetsnivåene mellom stimulansegenskapene til målet og distraktorene. Jo mer lik målet er til distraktoren, desto vanskeligere er det for observatøren å unngå fangst.
Parietallobens Rolle I Topp-Ned Og Bunn-Opp-Utvalg: Neuroimaging
Ulike neuroimaging teknikker gitt sterke bevis for involvering av parietal cortex i top-down og bottom-up orientering, med bevis anmeldt mye andre steder (Corbetta Og Shulman, 2002, 2011; Behrmann et al., 2004). Det har blitt påvist at områder som oftest aktiveres etter topp-ned-signaler for å delta på bestemte steder, funksjoner eller gjenstander, befinner seg langs dorsale deler av parietal cortex. Slike områder inkluderer inferior parietal lobule (IPL), dorsomediale regioner referert til som superior parietal lobule (SPL), samt flere mediale regioner langs precuneus gyrus (Yantis et al., 2002; Giesbrecht et al., 2003; Liu Et al., 2003; Yantis og Serences, 2003; Figur 1). Flere top-down oppgaver har vært viser å kunne engasjere dorsal regioner av parietal cortex, nemlig de som involverer romlig (Kastner et al., 1999; Corbetta et al., 2000; Hopfinger et al., 2000; Shomstein Og Behrmann, 2006; Chiu og Yantis, 2009; Greenberg et al., 2010) så vel som ikke-romlige skift av oppmerksomhet (Giesbrecht et al., 2003; Yantis og Serences, 2003; Shomstein Og Yantis, 2004b, 2006; Tamber-Rosenau et al., 2011).
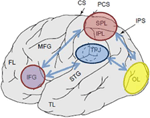
Figur 1. Skjematisk skildring av relevante anatomiske landemerker projisert på den laterale overflaten av den menneskelige hjerne. Superior parietal lobule (SPL) og inferior parietal lobule (IPL) er regioner innenfor den dorsale delen av parietal cortex subserving top-down attentional orientering. Temporo-parietal junction (tpj) er en region i den ventrale parietale cortex som subserverer bunn-opp oppmerksomhetsorientering. BÅDE SPL OG TPJ antas å fremkalle kontrollsignaler som er ansvarlige for etterfølgende oppmerksomhetsmoduleringer observert over sensoriske regioner, i dette tilfellet modulerende (merket med mørkeblå piler) visuelt fremkalt aktivitet i oksipitalloben (OL). I tillegg antas områder langs inferior frontal gyrus (IFG) og inferior frontal junction (IFJ) å tjene som konvergensområder for stimulusdrevet og topp-ned oppmerksomhetskontroll (merket med lyseblå toveis piler).
i en typisk oppgave som tar sikte på å engasjere top-down attentional allocation, blir individer vist to rapid serial visual presentation (rsvp) strømmer plassert perifert og blir først instruert til å overvåke en strøm for en cue (f.eks. et siffer blant strømmen av bokstaver). Identiteten til køen indikerer om motivet må opprettholde oppmerksomhet på gjeldende strøm eller skifte oppmerksomhet til den andre strømmen (Yantis et al.( 2002; Yantis og Serences, 2003). To store funn er observert i slike paradigmer. Den første har å gjøre med økt aktivering i de sensoriske områdene som representerer øyeblikket som deltok (f.eks. økt aktivitet i venstre primære visuelle regioner når den høyre RSVP-strømmen er deltatt). Dette funnet gir fast bevis på at deltakerne deltar på et bestemt sted, og at oppmerksomheten modulerer styrken av sensorisk respons (Se Figur 1; Moran Og Desimone, 1985; O ‘ Craven et al., 1997). Det andre funnet har å gjøre med observasjonen at dorsale regioner i parietalloben selektivt aktiveres av skift av topp-ned-oppmerksomhet. DET er observert AT SPL / IPL timecourse av aktivitet er forbigående i naturen som tyder på at dette området av parietal cortex er kilden til en kort attentional kontroll signal for å skifte oppmerksomme tilstander i en top-down måte (Yantis et al., 2002).
Flere fMRI-studier har dokumentert at nedenfra og opp oppmerksomhetsfangst, mediert av stimulus salience og / eller relevans, subserveres av temporo-parietal veikryss (Tpj; Figur 1). For eksempel, når fag ivareta og overvåke en endring i enten en visuell eller auditiv stimulus, presentert samtidig, aktivering AV tpj regioner av parietallappen er forbedret. I tillegg til den tilsynelatende følsomheten for relevante stimuli, aktiveres TPJ også som svar på potensielt nye (uventede eller sjeldne) hendelser når en organisme er engasjert i en nøytral atferdskontekst eller når den er engasjert i en oppgave (Marois et al., 2000; Nedar et al., 2002; Serences et al., 2005; Corbetta et al., 2008; Asplund et al., 2010; Diquattro Og Geng, 2011; Geng Og Mangun, 2011). Denne aktiveringen skjer uavhengig av modaliteten (auditiv, taktil og visuell) der inngangen leveres, noe som reflekterer tpjs multisensoriske natur (men se Downar et al., 2001).
i en typisk oppgave som undersøker nevrale mekanismen for bottom-up attentional capture, presenteres deltakerne med EN RSVP-strøm av elementer i midten av skjermen og blir bedt om å identifisere et forhåndsdefinert mål (f.eks. identifisere rødt brev presentert i en rsvp-strøm av hvite ikke-mål). En del av forsøkene inneholder en oppgave-irrelevant fremtredende distractor presentert på ulike tidsintervaller før utbruddet av målet, mens andre forsøk inneholder bare den fremtredende distractor (dvs.uten målet). «Target-distractor» forsøk brukes for å analysere omfanget av fangst, viser at oppgaven irrelevant distractor er faktisk fremtredende og dermed gir en reduksjon i mål nøyaktighet. «Distractor-in-isolation» – forsøkene brukes til videre analyser, siden slike forsøk tillater undersøkelse av aktivitet fremkalt til den fremtredende distraktoren uten forurensning fra målrelaterte prosesser. Flere viktige funn kommer fra slike paradigmer. For det første, når distraktorer er romlig skilt fra målstedet, blir fangstdraaktorer ledsaget av økt kortikal aktivitet i tilsvarende regioner i sensorisk cortex (f.eks. retinotopisk organisert visuell cortex; se Figur 1). Slike resultater gir sterke bevis på at under fangst blir romlig oppmerksomhet faktisk fanget til den romlige plasseringen okkupert av distraktoren (Serences et al., 2005). For det andre aktiveres ventrale områder av parietal cortex, hovedsakelig INNENFOR TPJ selektivt av bottom-up, ufrivillig, skift av oppmerksomhet. Akkurat som aktivitet innenfor SPL for top-down orientering, timecourse av aktivitet observert OVER TPJ er forbigående i naturen som tyder på at denne regionen er kilden til en kort attentional kontroll signal for å skifte oppmerksomhet i en bottom-up måte.Det skal bemerkes at mens denne gjennomgangen er fokusert på å adressere kognitive funksjoner i den bakre parietale cortexen, rekrutteres også andre regioner, spesielt de i frontal cortex for topp-ned og bunn-opp oppmerksomhetsallokering. Slike regioner inkluderer ventral frontal cortex (VFC), frontal eye fields (FEF), inferior frontal junction (IFJ) og inferior frontal gyrus (IFG; Corbetta and Shulman, 2002, 2011; Serences et al., 2005; Asplund et al.(2010; Diquattro Og Geng, 2011).
Parietallobens Rolle I Topp-Ned Og Bunn-Opp-Utvalg: Nevropsykologi
historisk forskere støttet seg kritisk på nevropsykologiske studier av pasienter med hemispatial forsømmelse (en lidelse av romlig tildeling av oppmerksomhet til venstre hemi-plass) for å få innsikt i kognitive funksjoner knyttet til parietalloben. I den klassiske nevropsykologiske litteraturen ble parietal cortex som helhet generelt ansett som det primære lesionsstedet for hemispatial forsømmelse. Denne visningen, utarbeidet i detalj av tidlige forskere (Critchley, 1953; McFire Og Zangwill, 1960; Piercy, 1964) anerkjente tydelig sammenhengen mellom parietal lesjon og den påfølgende forsømmelsen. Dette perspektivet ble i stor grad holdt gjennom 1980-tallet da Posner og kolleger (1984) brukte det skjulte visuospatiale cueing-paradigmet for å vise at skade på parietalloben gir et underskudd i» frakobling » – operasjonen (trekker oppmerksomheten fra ett sted og skifter den til en annen) når målet er kontralateralt til lesjonen. Til tross for dette store fremskrittet i å forstå det nevrale grunnlaget for oppmerksomhet og spesielt» løsne » rollen av parietal cortex, antar deres funn et enkelt kortikalt sted (parietal cortex) og en enkelt funksjonell evne («løsne»). I motsetning til denne mer monolitiske tilnærmingen til hjernen (parietal cortex) og atferd (attentional disengagement), tyder nyere atferds-og neuroimaging-arbeid (gjennomgått ovenfor og andre steder) på at både den kortikale regionen og tilhørende oppmerksomhetsadferd kan deles inn i kvalitativt forskjellige profiler.Gitt segregering av de kortikale nettverkene i topp-ned og bunn-opp prosesser, er en åpenbar prediksjon at skade på overlegne deler av parietallobulen (subsuming SPL) skal gi et underskudd i målrettet oppmerksomhetsorientering, mens skade på de dårligere delene av parietallobulen (subsuming TPJ) vil resultere i et underskudd forbundet med stimulusdrevet oppmerksomhetsopptak. I den grad disse hjerneadferdskorrespondansene har blitt utforsket i den nevropsykologiske litteraturen, er denne prediksjonen ikke åpenbart opprettholdt. For eksempel er kliniske symptomer på hemispatial forsømmelse sterkt forbundet med skade på de dårligere delene av parietalloben, som inkluderer TPJ, i stedet for overlegne deler SOM SPL (Friedrich et al ., 1998; Shomstein et al.(2010; Corbetta Og Shulman, 2011). Dette er noe i strid med neuroimaging litteraturen, noe som tyder på at ROLLEN TIL TPJ er i fangst av oppmerksomhet, snarere enn i frivillig orientering av oppmerksomhet, domenet der forsømmelse pasienter synes å ha mest problemer. For å komplisere saken ytterligere, har det blitt bemerket at lesjoner som involverer SPL utelukkende, bare sjelden gir klinisk bevis på forsømmelse (Vallar Og Perani, 1986). En annen fersk studie med pasienter med lesjoner sentrert primært OVER TPJ OG STG men bevart SPL, Corbetta et al. (2005) viste at romlig forsømmelse, så vel som gjenoppretting, var knyttet til restaurering av aktivitet i både ventral temporo-parietal og dorsal parietal regioner(se Corbetta and Shulman, 2011 for en gjennomgang). Mens det er interessant og spennende i sine konklusjoner, skiller denne siste studien ikke det relative bidraget av dorsale og ventrale veier til forskjellige typer oppmerksomhet, siden pasientene bare ble testet på En variant Av Posner covert romlig oppmerksomhet cuing oppgave, oppgave som antas å engasjere både topp-ned og bunn-opp oppmerksomhetsorientering.for å skille mellom måldrevet oppmerksomhetskontroll og fremtredende oppmerksomhetsfangst og å undersøke kartleggingen på henholdsvis SPL og TPJ, vedtok en nylig studie to atferdsparadigmer, hver rettet mot en av disse formene for oppmerksomhet (Shomstein et al ., 2010). For å undersøke integriteten til top-down attentional orientering hos pasientene, ble det brukt en topp-down-oppgave som krever at deltakerne skifter romlig oppmerksomhet mellom romlig adskilte RSVP-strømmer (en oppgave som har blitt brukt til å demonstrere SPL-aktivering i fMRI-studier (Yantis et al ., 2002)). På samme måte, for å undersøke pasientens nedre oppmerksomhetsorienterende evner, en variant Av Folk et al. (2002) kontingent capture paradigme ble ansatt der deltakerne oppdaget mål som dukket opp ved fiksering mens oppgaverelevante farge enkeltfødte ble blinket i periferien. I hvilken grad oppgaverelevante distraktorer forstyrrer den sentrale deteksjonsoppgaven, ble deretter brukt som et mål for bunn-opp oppmerksomhetsfangst (Bacon Og Egeth, 1994; Folk et al., 2002).
spådommene var som følger: pasienter med lesjoner til overlegne deler av parietallappen (påvirker SPL) bør være svekket i topp-ned oppmerksomhetsorienteringsoppgaven (med bevart ytelse på fangstoppgaven), mens pasienter med lesjoner til de dårligere delene av parietallappen (påvirker TPJ) bør være svekket på fangstoppgaven (med spart ytelse på topp-ned-oppgaven). En dobbel dissosiasjon av dette skjemaet vitner ikke bare om de uavhengige komponentene av oppmerksomhet, men foreslår også at slike oppmerksomhetskomponenter medieres av uavhengige nevrale mekanismer. Åtte pasienter med visuo-romlig forsømmelse ble rekruttert til studien og fullførte to oppgaver, tapping enten stimulusdrevet eller målrettet oppmerksomhetsorientering. Basert på deres atferdsprofil ble pasientene sortert i grupper og deres lesjonsoverlapping ble utforsket (Figur 2a). Pasienter som viste vanskeligheter med målrettet oppmerksomhetsorientering, som kvantifisert av top-down attentional index (Figur 2b), presentert med lesion overlapping sentrert over overlegne deler av parietal lobule (subsuming SPL) med spart dårligere parietal lobule (TPJ). Pasienter med lesion overlapping sentrert over de dårligere delene av parietal lobule (subsuming TPJ), men spart SPL utført normalt på målrettet orienteringsoppgave, mens resterende immun mot attentional fangst (Figur 2c). Funnene fra denne studien tyder tydelig på AT SPL og TPJ er anatomiske regioner som nødvendigvis rekrutteres med henblikk på top-down og bottom-up orientering, og at skade PÅ SPL og TPJ fører til forstyrrelser i henholdsvis top-down og bottom-up orientering.
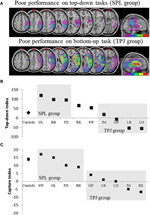
Figur 2. Resultatene av den nevropsykologiske studien rettet mot å undersøke det relative bidraget TIL SPL OG TPJ til topp-ned og bunn-opp orientering. (A) Lesion overlapper (lilla minimal overlapping; rød maksimal overlapping) for pasienter gruppert etter atferdsmessige underskudd i topp-ned oppmerksomhetsorientering, merket SPL-gruppen (topppanel); og pasienter gruppert etter atferdsmessige underskudd i bunn-opp orientering, merket TPJ-gruppen (nedre panel). (B) Atferdsmessig ytelse på topp-ned-oppgaven oppsummert med En «Topp-ned-Indeks» som kvantifiserer forskjeller mellom romlige topp-ned-skift fra venstre til høyre og omvendt. Kontroller og tpj lesioned-gruppen viser lignende effektivitet i å utføre romlige skift, mens pasienter med SPL-lesjoner viser redusert effektivitet. Gruppekontroll og individuelle pasientdata (merket med pasientinitialer) er plottet på abscissen. (C) «Capture index» er et mål på bottom-up oppmerksomhet og kvantifiserer i hvilken grad oppgave-irrelevante distraktorer fanger oppmerksomheten bort fra oppgaven. Kontroller og spl lesioned-gruppen viser lignende opptaksverdier, slik at begge gruppene fanges opp av aktivitetsrelevante distraktorer. TPJ lesioned group viser mye redusert fangstindeks (unnlatelse av å bli fanget). Merk at pasientene ble plassert i SPL-eller TPJ-gruppen basert på atferd, i stedet for basert på lesjonen, og dermed merk konsistensen som pasientene ender i den tilsvarende gruppen.
Interaksjon mellom Top-Down Og Bottom-Up Utvalg
selv om det er tilsynelatende en sterk sammenheng mellom målrettet orientering OG SPL og stimulus-drevet orientering OG TPJ, data Fra Shomstein et al. (2010) pasientstudie tyder på at disse to systemene ikke er helt uavhengige. Denne konklusjonen støttes av funnet at pasienter med SPL-skade viste et mønster av ytelse merket som » hyper capture.»I motsetning til kontroller, for hvem bare målfarget distraktor fanget oppmerksomhet (som fører til lavere målnøyaktighet), viste irrelevante fargede distraktorer seg også å være distraherende for pasienter med SPL-lesjon. I tillegg, mens for kontroller oppmerksomhet ble fanget av distraktorer bare når de gikk foran målets begynnelse, ble det for pasienter med SPL-lesjoner også tatt oppmerksomhet av distraktorer presentert samtidig med målet. Dette ytelsesmønsteret kan forklares av følgende rammeverk: SPL er ansvarlig for top-down veiledning av oppmerksomhet som inkluderer å bestemme aspekter av stimuli som er oppgave relevant (f. eks søk etter red target; Corbetta og Shulman, 2002; Serences et al., 2005). Dette oppmerksomhetssettet begrenser DERETTER TPJ, slik at fangst av oppmerksomhetsmekanisme som formidles AV TPJ, bare utløses av oppgavens relevante informasjon(f. eks. Fraværet AV SPL hindrer etablering av en oppgave relevant oppmerksomhet sett og dermed enhver stimulans, oppgave relevant eller ikke, anses viktig derfor fange oppmerksomhet (f. eks oppgave-irrelevant distraktor fange oppmerksomhet FOR SPL gruppen) ukritisk.
DET har blitt foreslått AT SPL og TPJ kunne samhandle på minst en av to mulige måter. DEN første muligheten er AT TPJ fungerer som et varslingssystem som oppdager atferdsrelevante stimuli, men mangler den høye romlige oppløsningen, og når en atferdsrelevant stimulus oppdages, blir den nøyaktige plasseringen levert AV SPL som lagrer romlige kart(Kastner et al ., 1999; Wojciulik Og Kanwisher, 1999; Bisley Og Goldberg, 2003; Silver et al., 2005). En relatert hypotetisk mulighet er at fangstmekanismen (som inkluderer TPJ) fungerer som en bryter for pågående kognitiv aktivitet når en atferdsrelevant stimulus presenteres (Corbetta og Shulman, 2002, 2011). «Hyper-capture» aktivitetsmønsteret observert hos pasienter med bevart TPJ, men lesioned SPL gir ytterligere bevis for hypotesen om at ser TPJ som å utstede et kontrollsignal som avslutter oppgaven ved hånden og dermed tjener som en bryter (Corbetta Og Shulman, 2002; Serences et al., 2005). Andre nyere neuroimaging studier som bruker ulike paradigmer har gitt ytterligere bevis for et interaktivt forhold mellom topp-ned og bunn-opp oppmerksom orientering, og senere for forholdet MELLOM SPL og TPJ (Serences et al., 2005; Asplund et al., 2010; Diquattro Og Geng, 2011).mens bevisene for en interaksjon mellom de to oppmerksomhetssystemene og DE TO oppmerksomhetssubstratene (SPL og TPJ) er sterke, er det uklart om denne interaksjonen er direkte mellom SPL og TPJ, eller om den oppnås gjennom andre mellomliggende regioner. Som nevnt tidligere, top-down og bottom-up attentional orienteringsnettverk engasjere ulike regioner innenfor frontal cortex, og det er derfor rimelig å hypotese at konvergensen mellom de to systemene kan oppnås via frontal lobe. To nyere studier undersøker samspillet mellom top-down og bottom-up attentional selection gitt bevis FOR IFJ OG IFG som mulige områder av konvergens mellom stimulus-drevet og målrettet seleksjon (Asplund et al.(2010; Diquattro Og Geng, 2011). IFJ og IFG synes å være ideelle kandidater for slik interaksjon gitt deres generelle engasjement i oppmerksomhet og kognitiv kontroll, samt sitt engasjement i både romlig og ikke-romlig utvalg (Koechlin et al., 2003; Brass et al., 2005).
Parietallobens Rolle I Topp-Ned Og Bunn-Opp-Utvalg: Fysiologi
mens vekten av denne anmeldelsen hovedsakelig er lagt på menneskelige studier, har en stor mengde kunnskap om involvering av parietal cortex i oppmerksom orientering blitt hentet fra apefysiologiske undersøkelser (se nylig gjennomgang Av Bisley og Goldberg, 2010). Men når det gjelder å undersøke de relative bidragene fra ulike regioner i parietal cortex til topp-ned og bunn-opp oppmerksomhet orientering, faller monkey fysiologi litteratur kort. Den primære årsaken til dette er at i monkey cortex ser det ikke ut til å være bevis for samme segregering av top-down og bottom-up kontroll. I stedet er lateral intraparietal area (LIP) opprinnelig antatt å være involvert i saccade planlegging (Gnadt Og Andersen, 1988) involvert i visuell oppmerksomhet og fungerer som et prioriteringskart der eksterne stimuli er representert i henhold til deres atferdsprioritet avledet på enten topp-ned eller bunn-opp måte (Colby og Goldberg, 1999; Bisley og Goldberg, 2003, 2010; Balan og Gottlieb, 2006; Ipata et al., 2006; Buschman Og Miller, 2007; Gottlieb Og Balan, 2010).
Konklusjon
selv om mye mindre er kjent om human parietal cortex enn for homolog ape cortex, nyere studier, ansette neuroimaging og nevropsykologiske metoder, har begynt å belyse stadig finkornet funksjonelle og strukturelle forskjeller. Denne gjennomgangen fokuserte på nyere neuroimaging og nevropsykologiske studier som belyser de kognitive rollene til dorsale og ventrale regioner av parietal cortex i topp-ned og bunn – opp oppmerksomhetsorientering, og på samspillet mellom de to oppmerksomhetsallokeringsmekanismer.
Interessekonflikt
forfatteren erklærer at forskningen ble utført i fravær av kommersielle eller økonomiske forhold som kan tolkes som en potensiell interessekonflikt.
Takk
dette arbeidet ble støttet Av National Institutes Of Health grant EY021644.Asplund, C. L., Todd, J. J., Snyder, A. P., Og Marois, R. (2010). En sentral rolle for den laterale prefrontale cortex i målrettet og stimulusdrevet oppmerksomhet. Nat. Neurosci. 13, 507–512.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Bacon, Wf, Og Egeth, H. E. (1994). Overstyre stimulus-drevet oppmerksomhetsfangst. Percept. Psykofyser. 55, 485–496.
Pubmed Abstract/Pubmed Full Text
Balan, P. F. Gottlieb, J. (2006). Integrering av eksogene innspill i en dynamisk salience kart avslørt av perturbing oppmerksomhet. J. Neurosci. 26, 9239–9249.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Behrmann, M., Geng, Jj, Og Shomstein, S. (2004). Parietal cortex og oppmerksomhet. Curr. Opin. Neurobiol. 14, 212–217.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Bisley, J. W., Og Goldberg, Me (2003). Neuronal aktivitet i det laterale intraparietale området og romlig oppmerksomhet. Vitenskap 299, 81-86.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Bisley, J. W., Og Goldberg, Me (2010). Oppmerksomhet, intensjon og prioritet i parietalloben. Annu. Pastor Neurosci. 33, 1–21.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Brass, M., Derrfuss, J., Forstmann, B., og von Cramon, D. Y. (2005). Rollen til det dårligere frontale veikryssområdet i kognitiv kontroll. Trender Cogn. Sci. 9, 314–316.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Buschman, T. J., Og Miller, E. K. (2007). Top-down versus bottom-up kontroll av oppmerksomhet i prefrontal og posterior parietal cortices. Vitenskap 315, 1860-1862.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Chiu, Yc, Og Yantis, S. (2009). En domeneuavhengig kilde til kognitiv kontroll for oppgavesett: skiftende romlig oppmerksomhet og bytte kategoriseringsregler. J. Neurosci. 29, 3930–3938.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Colby, C. L., Og Goldberg, Me (1999). Plass og oppmerksomhet i parietal cortex. Annu. Pastor Neurosci. 22, 319–349.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., Kincade, Jm, Ollinger, Jm, McAvoy, Mp, Og Shulman, Gl (2000). Frivillig orientering er dissosiert fra måldeteksjon i menneskelig bakre parietal cortex. Nat. Neurosci. 3, 292–297.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., Kincade, M. J., Lewis, C., Snyder, A. Z., Og Sapir, a. (2005). Nevrale grunnlag og gjenoppretting av romlige oppmerksomhetsunderskudd i romlig forsømmelse. Nat. Neurosci. 8, 1603–1610.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Corbetta, M., Patel, G., Og Shulman, G. L. (2008). Det omorienterende systemet i den menneskelige hjerne: fra miljø til sinnsteori. Neuron 58, 306-324.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Corbetta, M., Og Shulman, G. L. (2002). Kontroll av målrettet og stimulusdrevet oppmerksomhet i hjernen. Nat. Pastor Neurosci. 3, 201–215.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Corbetta, M., Og Shulman, G. L. (2011). Romlig forsømmelse og oppmerksomhet nettverk. Annu. Pastor Neurosci. 34, 569–599.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Critchley, M. (1953). Parietallobene. London, STORBRITANNIA: Hafner Press.
Pubmed Abstract/Pubmed Full Text
Diquattro, N. E., Og Geng, J. J. (2011). Kontekstuell kunnskap konfigurerer oppmerksomhetskontrollnettverk. J. Neurosci. 31, 18026–18035.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Downar, J., Crawley, Ap, Mikulis, Dj, Og Davis, K. d. (2001). Effekten av oppgaverelevans på kortikal respons på endringer i visuelle og auditive stimuli: en hendelsesrelatert fmri-studie. Neuroimage 14, 1256-1267.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Downar, J., Crawley, Ap, Mikulis, Dj, Og Davis, K. d. (2002). Et kortikalt nettverk som er følsomt for stimulans salience i en nøytral atferdskontekst på tvers av flere sensoriske modaliteter. J. Nevrofysiol. 87, 615–620.
Pubmed Abstract/Pubmed Full Text
Drummond, L., Og Shomstein, S. (2010). Objektbasert oppmerksomhet: endring eller usikkerhet? Aten. Percept. Psykofyser. 72, 1743–1755.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Egeth, H. E., Og Yantis, S. (1997). Visuell oppmerksomhet: kontroll, representasjon og tidskurs. Annu. Pastor Psychol. 48, 269–297.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Eriksen, C. W., Og Hoffman, J. E. (1972). Tidsmessige og romlige egenskaper ved selektiv koding fra visuelle skjermer. Percept. Psykofyser. 12, 201–204.
Folk, C. L., Leber, A. B., Og Egeth, H. E. (2002). Fikk deg til å blunke! Betinget attentional fangst produserer en romlig blink. Percept. Psykofyser. 64, 741–753.
Pubmed Abstract/Pubmed Full Text
Folk, C. L., Remington, R. W., Og Johnston, J. C. (1992). Ufrivillig skjult orientering er betinget av oppmerksomhetskontrollinnstillinger. J. Exp. Psychol. Hum. Percept. Utføre. 18, 1030–1044.
Pubmed Abstract/Pubmed Full Text
Friedrich, F. J., Egly, R., Rafal, R. D., Og Beck, D. (1998). Romlig oppmerksomhet underskudd hos mennesker: en sammenligning av overlegen parietal og temporal-parietal krysset lesjoner. Neuropsykologi 12, 193-207.
Pubmed Abstract/Pubmed Full Text
Geng, J. J., Og Behrmann, M. (2002). Sannsynlighet cuing av målplassering letter visuelt søk implisitt hos normale deltakere og pasienter med hemispatial forsømmelse. Psychol. sci. 13, 520–525.
Pubmed Abstract/Pubmed Full Text
Geng, J. J., Og Behrmann, M. (2005). Romlig sannsynlighet som en oppmerksomhetssignal i visuelt søk. Percept. Psykofyser. 67, 1252–1268.
Pubmed Abstract/Pubmed Full Text
Geng, J. J., And Mangun, G. R. (2011). Høyre temporoparietal veikryss aktivering av en fremtredende kontekstuell kø letter måldiskriminering. Neuroimage 54, 594-601.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Gibson, Bs, Og Kelsey, E. M. (1998). Stimulus-drevet attentional capture er betinget av attentional sett for displaywide visuelle funksjoner. J. Exp. Psychol. Hum. Percept. Utføre. 24, 699–706.
Pubmed Abstract/Pubmed Full Text
Giesbrecht, B., Woldorff, Mg, Song, A. W., Og Mangun, G. R. (2003). Nevrale mekanismer av top-down kontroll under romlig og har oppmerksomhet. Neuroimage 19, 496-512.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Gnadt, J. W., And Andersen, R. A. (1988). Minne relatert motor planlegging aktivitet i bakre parietal cortex av macaque. Exp. Hjernereses70, 216-220.
Pubmed Abstract/Pubmed Full Text
Gottlieb, J., Og Balan, P. (2010). Oppmerksomhet som en beslutning i informasjonsområdet. Trender Cogn. Sci. 14, 240–248.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Greenberg, A. S., Esterman, M., Wilson, D., Serences, Jt, Og Yantis, S. (2010). Kontroll av romlig og funksjonsbasert oppmerksomhet i frontoparietal cortex. J. Neurosci. 30, 14330–14339.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Hopfinger, J. B., Buonocore, M. H., Og Mangun, G. R. (2000). Nevrale mekanismer av topp-ned oppmerksomhetskontroll. Nat. Neurosci. 3, 284–291.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Ipata, A. E.( 2006), Gee, A. L., Gottlieb, J., Bisley, J. W., Og Goldberg, Me. LEPPE svar på en popout stimulus reduseres hvis det er åpenlyst ignorert. Nat. Neurosci. 9, 1071–1076.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Johnston, W. A., And Dark, V. J. (1986). Selektiv Oppmerksomhet. Annu. Pastor Psychol. 37, 43–75.I 1995 ble Det opprettet En egen organisasjon. Avveininger mellom attentional effekter av romlige signaler og brå onsets. Percept. Psykofyser. 57, 333–342.
Pubmed Abstract/Pubmed Full Text
Kastner, S., Pinsk, M. A., De Weerd, P., Desimone, R., Og Ungerleider, L. G. (1999). Økt aktivitet i menneskelig visuell cortex under rettet oppmerksomhet i fravær av visuell stimulering. Neuron 22, 751-761.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Koechlin, E., Ody, C., Og Kouneiher, F. (2003). Arkitekturen av kognitiv kontroll i den menneskelige prefrontale cortex. Vitenskap 302, 1181-1185.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Koshino, H. Warner, C. B. Og Juola, J. F. (1992). Relativ effekt av sentrale, perifere og brå signaler i visuell oppmerksomhet. Q. J. Exp. Psychol. En 45, 609-631.
Pubmed Abstract/Pubmed Full Text
Liu, T., Slotnick, S. D., Serences, J. T., Og Yantis, S. (2003). Kortikale mekanismer av funksjonsbasert oppmerksomhetskontroll. Cereb. Cortex 13, 1334-1343.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Marois, R., Leung, H. C., Og Gore, J. C. (2000). En stimulus-drevet tilnærming til objekt identitet og plassering behandling i den menneskelige hjerne. Neuron 25, 717-728.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
McFire, J., And Zangwill, O. L. (1960). Visuo-konstruktive funksjonshemninger forbundet med lesjoner i venstre hjernehalvdel. Hjerne 82, 243-260.
Moore, Cm, Og Egeth, H. (1998). Hvordan påvirker funksjonsbasert oppmerksomhet visuell behandling? J. Exp. Psychol. Hum. Percept. Utføre. 24, 1296–1310.
Pubmed Abstract/Pubmed Full Text
Moran, J. Og Desimone, R. (1985). Selektiv oppmerksomhet gates visuell behandling i extrastriate cortex. Vitenskap 229, 782-784.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
O ‘ Craven, K., Rosen, B. R., Kwong, K. K., Treisman, A., Og Savoy, R. L. (1997). Frivillig oppmerksomhet modulerer fMRI aktivitet i menneskelig MT-MST. Neuron 18, 591-598.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Piercy, M. (1964). Effektene av cerebrale lesjoner på intellektuell funksjon: en gjennomgang av dagens forskningstrender. Br. J. Psykiatri 110, 310-352.
Pubmed Abstract/Pubmed Full Text
Posner, M. I. (1980). Orientering av oppmerksomhet. Q. J. Exp. Psychol. 32, 3–25.
Pubmed Abstract/Pubmed Full Text
Posner, M. I., Snyder, C. R., Og Davidson, B. J. (1980). Oppmerksomhet og påvisning av signaler. J. Exp. Psychol. 109, 160–174.
Pubmed Abstract/Pubmed Full Text
Posner, M. I., Walker, J. A., Friedrich, F. J., Og Rafal, R. D. (1984). Effekter av parietal skade på skjult orientering av oppmerksomhet. J. Neurosci. 4, 1863–1874.
Pubmed Abstract/Pubmed Full Text
Serences, J. T., Shomstein, S., Leber, A. B., Golay, X., Egeth, H. E. og Yantis, S. (2005). Koordinering av frivillig og stimulusdrevet oppmerksomhetskontroll i menneskelig cortex. Psychol. Sci. 16, 114–122.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Shaw, M. L. (1978). En kapasitetstildelingsmodell for reaksjonstid. J. Exp. Psychol. Hum. Percept. Utføre. 4, 586–598.
Shomstein, S., Og Behrmann, M. (2006). Kortikale systemer som formidler visuell oppmerksomhet til både objekter og romlige steder. Proc. Natl. Acad. Sci. U. S. A. 103, 11387-11392.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Shomstein, S., Lee, J., Og Behrmann, M. (2010). Top-down og bottom-up attentional guidance: undersøker rollen til dorsal og ventral parietal cortices. Exp. Brain Res. 206, 197-208.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Shomstein, S., Og Yantis, S. (2004a). Konfigural og kontekstuell prioritering i objektbasert oppmerksomhet. Psychon. Bull. Åp 11, 247-253.
Pubmed Abstract/Pubmed Full Text
Shomstein, s., Og Yantis, S. (2004b). Kontroll av oppmerksomhet skifter mellom syn og audition i menneskelig cortex. J. Neurosci. 24, 10702–10706.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Shomstein, S., Og Yantis, S. (2006). Parietal cortex formidler frivillig kontroll av romlig og ikke-spatial auditiv oppmerksomhet. J. Neurosci. 26, 435–439.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Sølv, M. A., Ress, D. Og Heeger, D. J. (2005). Topografiske kart over visuell romlig oppmerksomhet i menneskelig parietal cortex. J. Nevrofysiol. 94, 1358–1371.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Tamber-Rosenau, B. J., Esterman, M., Chiu, Y.-C., Og Yantis, S. (2011). Kortikale mekanismer for kognitiv kontroll for å skifte oppmerksomhet i visjon og arbeidsminne. J. Cogn. Neurosci. 23, 2905–2919.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Theeuwes, J. (1991). Eksogen og endogen kontroll av oppmerksomhet: effekten av visuelle onsets og offsets. Percept. Psykofyser. 49, 83–90.
Pubmed Abstract/Pubmed Full Text
Vallar, G., Og Perani, D. (1986). Anatomien til ensidig forsømmelse etter høyre hjernehalvdel slaglesjoner. En klinisk / CT-skanning korrelasjonsstudie i man. Neuropsykologi 24, 609-622.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Wojciulik, E., Og Kanwisher, N. (1999). Generaliteten av parietal involvering i visuell oppmerksomhet. Neuron 23, 747-764.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Yantis, S., Og Jonides, J. (1984). Brå visuelle onsets og selektiv oppmerksomhet: bevis fra visuelt søk. J. Exp. Psychol. Hum. Percept. Utføre. 10, 601–621.
Pubmed Abstract/Pubmed Full Text
Yantis, S., Og Jonides, J. (1990). Brå visuelle onsets og selektiv oppmerksomhet: frivillig versus automatisk tildeling. J. Exp. Psychol. Hum. Percept. Utføre. 16, 121–134.
Pubmed Abstract/Pubmed Full Text
Yantis, S., Schwarzbach, J., Serences, J. T., Carlson, R. L., Steinmetz, M. A., Pekar, J. J., Og Courtney, S. M. (2002). Transient neural aktivitet i human parietal cortex under romlig oppmerksomhet skift. Nat. Neurosci. 5, 995–1002.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Yantis, S., and Serences, J. T. (2003). Kortikale mekanismer for rombasert og objektbasert oppmerksomhetskontroll. Curr. Opin. Neurobiol. 13, 187–193.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text