Kromatin er et kompleks av makromolekyler som består AV DNA, RNA og protein, som finnes inne i kjernen av eukaryote celler. Kromatin finnes i to former: heterochromatin (kondensert) og eukromatin (utvidet). De primære proteinkomponentene i kromatin er histoner som bidrar til å organisere DNA i «perle-lignende» strukturer kalt nukleosomer ved å gi en base SOM DNA kan vikles rundt. Et nukleosom består av 147 basepar DNA som er viklet rundt et sett med 8 histoner kalt en oktomer. Nukleosomet kan videre foldes for å produsere kromatinfiberen. Kromatinfibre spoles og kondenseres for å danne kromosomer. Kromatin gjør det mulig for EN rekke celleprosesser å skje, inkludert DNA-replikasjon, transkripsjon, DNA-reparasjon, genetisk rekombinasjon og celledeling.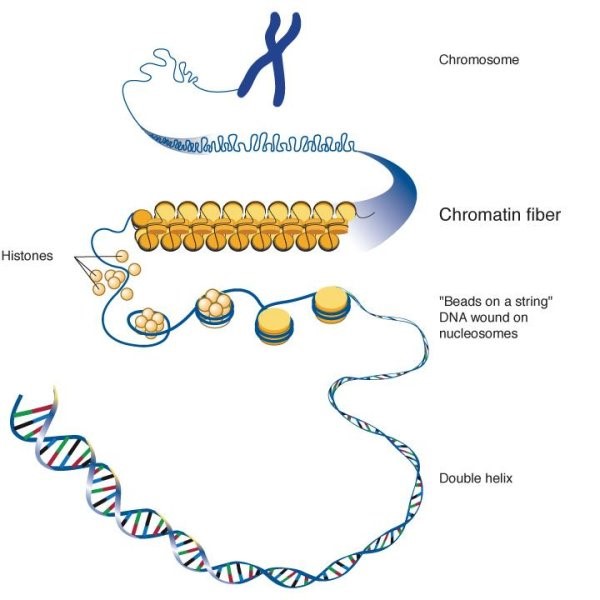
Kromatin, Kromosomer og Kromatider
folk forveksler ofte disse tre begrepene: kromatin, kromosom og kromatid. Mens ALLE disse tre strukturene er sammensatt AV DNA og proteiner i kjernen, er hver unikt definert.
som nevnt ovenfor består kromatin AV DNA og histoner som er pakket inn i tynne, trevlete fibre. Kromatinet gjennomgår ytterligere kondensering for å danne kromosomet. Så kromatin er en lavere REKKEFØLGE AV DNA-organisasjon, mens kromosomer er den høyere rekkefølgen AV DNA-organisasjon.
Kromosomer er enkeltstrengede grupperinger av kondensert kromatin. Under celledeling prosesser av mitose og meiose, kromosomer replikere for å sikre at hver ny datter celle får riktig antall kromosomer. Et duplisert kromosom er dobbeltstrenget og har den kjente x-formen. De to strengene er identiske og koblet til en sentral region kalt centromere.
en kromatid er en av de to trådene i et replikert kromosom. Kromatider forbundet med en centromere kalles søsterkromatider. På slutten av celledeling separerer søsterkromatider og blir datterkromosomer i de nyopprettede dattercellene.
Funksjonen Av Kromatin
DNA-Emballasje
dette er den mest grunnleggende funksjonen til kromatin: komprimering av lange DNA-tråder.LENGDEN PÅ DNA i kjernen er langt større enn størrelsen på rommet der den er lagret. FOR å passe inn i DETTE rommet MÅ DNA kondenseres på noen måte. Pakningsforhold brukes til å beskrive i hvilken grad DNA kondenseres. FOR å oppnå det totale pakningsforholdet pakkes IKKE DNA direkte inn i strukturen av kromatin. I stedet inneholder den flere hierarkier av organisasjon.
det første nivået av pakking oppnås ved vikling AV DNA rundt nukleosomet, noe som gir et pakningsforhold på ca.6. Denne strukturen er invariant i både eukromatin og heterochromatin av alle kromosomer. Det andre nivået av pakking er innpakning av perler i en 30 nm fiber som finnes i både interfasekromatin og mitotiske kromosomer. Denne strukturen øker pakningsforholdet til ca 40. Den endelige emballasjen oppstår når fiberen er organisert i løkker, stillaser og domener som gir et endelig pakningsforhold på ca.1000 i interfasekromatin og ca. 10.000 i mitotiske kromosomer.Transkripsjon Er en Prosess der den genetiske informasjonen lagret I DNA leses av proteiner og deretter transkriberes TIL RNA, og RNA vil senere bli oversatt til funksjonelle proteiner. Hvis kromatinet blir styrket og begrenser tilgangen til leseproteinene, skjer det ingen transkripsjon. Euchromatin, en utvidet type kromatin, kan gjennomføre transkripsjonsprosessen. Mens heterochromatin, den kondenserte typen kromatin, er pakket for tett FOR AT DNA kan leses av proteiner.
Svingninger mellom åpen og lukket kromatin kan bidra til diskontinuitet av transkripsjon, eller transkripsjonelle sprengning. Andre faktorer kan trolig være involvert, for eksempel forening og dissosiasjon av transkripsjonsfaktorkomplekser med kromatin. Fenomenet, i motsetning til enkle probabilistiske modeller for transkripsjon, kan utgjøre den høye variabiliteten i genuttrykk som forekommer mellom celler i isogen populasjon
Kromatin og DNA-Reparasjon
emballasjen AV DNA i kromatin presenterer en barriere for ALLE DNA-baserte prosesser. På grunn av det høye dynamiske arrangementet av proteiner og DNA, kan kromatin lett endre form og struktur. Kromatinavslapping skjer raskt på STEDET FOR DNA-skade, noe som gjør at reparasjonsproteinene kan binde SEG TIL DNA og reparere DET.
Referanse:
1. Comings D E. Strukturen og funksjonen til kromatin . Fremskritt i menneskelig genetikk. Springer USA, 1972: 237-431.
2. Widom J. Struktur, dynamikk og funksjon av kromatin in vitro . Årlig gjennomgang av biofysikk og biomolekylær struktur, 1998, 27(1): 285-327.
3. Mercer T R, Mattick J S. Struktur Og funksjon av lange ikke-kodende Rna i epigenetisk regulering . Natur strukturelle & molekylærbiologi, 2013, 20(3): 300-307.