introducere
interacțiunea reușită cu mediul nostru senzorial necesită un echilibru complicat de două mecanisme de selecție atențională-cea de sus în jos și de jos în sus. Îndreptându-vă spre culoarul de produse al supermarketului dvs. local, cu scopul de a ridica câteva ingrediente necesare pentru salata de mango, angajează implementarea unui sistem atențional voluntar orientat spre obiectiv sau de sus în jos, astfel încât să căutați în mod activ toate ingredientele necesare printre multitudinea de alegeri de produse. Cu toate acestea, dacă auziți un sunet al unui telefon mobil, cel mai probabil vă va capta atenția și vă va întrerupe căutarea. O astfel de întrerupere are loc într-un mod de jos în sus sau stimulat, prin care o simplă proeminență a stimulului, faptul că inelul este diferit de alte sunete din mediul dvs., îl consideră demn de selecție. Scenariul descris subliniază importanța selecției orientate spre obiective și stimulate pentru comportament și indică un echilibru fin care trebuie să existe între cele două sisteme atenționale pentru a preveni „viziunea tunelului” pe de o parte și incapacitatea completă de a se concentra pe cealaltă.
selecție de sus în jos și de Jos în sus: Comportamentul
câteva decenii de cercetare comportamentală au fost dedicate demonstrării faptului că distribuția atenției poate fi controlată de intențiile observatorului, precum și de proeminența stimulului fizic. O mare parte din dovezile comportamentale pentru alocarea atențională de sus în jos și de jos în sus au fost revizuite pe larg în altă parte (Johnston și Dark, 1986; Egeth și Yantis, 1997). Pentru a rezuma, studiile care demonstrează efectele controlului atențional de sus în jos arată că atenția poate fi alocată cu succes locațiilor spațiale, caracteristicilor, obiectelor etc., în urma prezenței indiciilor exogene sau endogene (Eriksen și Hoffman, 1972; Posner, 1980; Posner și colab., 1980) sau așteptări stabilite fie de cunoștințe anterioare, fie de contingențe ale stimulului (Shaw, 1978; Moore și Egeth, 1998; Geng și Behrmann, 2002, 2005; Shomstein și Yantis, 2004a; Drummond și Shomstein, 2010). Dovezile care susțin alocarea atențională de jos în sus s-au bazat pe diferite paradigme de captare atențională, în care participanții sunt angajați într-o căutare de sus în jos și atenția lor este deviată către stimulii irelevanți ai sarcinii, demonstrând că atenția este captată de singletons feature (element unic; Yantis și Jonides, 1990; Theeuwes, 1991; Folk și colab., 2002) și onsets abrupte (Yantis și Jonides, 1984; Theeuwes, 1991; Koshino și colab., 1992; Juola și colab., 1995).în timp ce majoritatea studiilor timpurii s-au concentrat pe demonstrarea dovezilor pentru selecția atențională de sus în jos și de jos în sus, cele mai recente studii și-au îndreptat atenția spre examinarea modului în care interacționează cele două sisteme de selecție atențională. Această linie de investigație este alimentată de observații conform cărora, pentru a selecta în mod eficient informații relevante pentru sarcină (de exemplu, ingrediente pentru salată), trebuie să inhibăm în mod activ informațiile irelevante despre sarcină, care altfel ar distrage atenția de la sarcina la îndemână. Reversul acestei logici, este că mai puțin unul este axat pe informații legate de sarcină mai mult de captare va rezulta. S-a demonstrat experimental că starea atențională a Observatorului prezice ce tip de informație și în ce măsură va capta în cele din urmă atenția (Folk și colab., 1992, 2002; Bacon și Egeth, 1994; Gibson și Kelsey, 1998). De exemplu, Folk și colab. (2002) a arătat că atunci când caută o literă roșie, un observator va fi mai ușor capturat de un stimul irelevant în periferie dacă acel stimul este roșu sau se potrivește într-un fel cu șablonul țintă. Deoarece setările de control de sus în jos ale observatorului sunt setate pentru a căuta o caracteristică roșie, orice stimul care este roșu este probabil să capteze atenția și să interfereze cu controlul de sus în jos. Astfel, cu o sarcină de captare, strategiile de căutare atențională pot fi distinse unele de altele prin modificarea nivelurilor de similitudine dintre proprietățile stimulului țintei și distractorii. Cu cât ținta este mai asemănătoare cu distractorul, cu atât este mai dificil pentru observator să evite capturarea.
rolul lobului Parietal în selecția de sus în jos și de Jos în sus: Neuroimagistica
diverse tehnici de neuroimagistică au furnizat dovezi puternice pentru implicarea cortexului parietal în orientarea de sus în jos și de jos în sus, dovezile fiind revizuite extensiv în altă parte (Corbetta și Shulman, 2002, 2011; Behrmann și colab., 2004). S-a demonstrat că zonele activate cel mai frecvent după indicii de sus în jos pentru a participa la anumite locații, caracteristici sau obiecte sunt situate de-a lungul părților dorsale ale cortexului parietal. Astfel de zone includ lobul parietal inferior (IPL), regiunile dorsomediale denumite lobul parietal superior (SPL), precum și mai multe regiuni mediale de-a lungul girusului precuneus (Yantis și colab., 2002; Giesbrecht și colab., 2003; Liu și colab., 2003; Yantis și Serences, 2003; Figura 1). Mai multe sarcini de sus în jos au fost spectacole pentru a angaja cu succes regiunile dorsale ale cortexului parietal, și anume cele care implică spațiu (Kastner și colab., 1999; Corbetta și colab., 2000; Hopfinger și colab., 2000; Shomstein și Behrmann, 2006; Chiu și Yantis, 2009; Greenberg și colab., 2010), precum și schimbări de atenție non-spațiale (Giesbrecht și colab., 2003; Yantis și Serences, 2003; Shomstein și Yantis, 2004b, 2006; Tamber-Rosenau și colab., 2011).
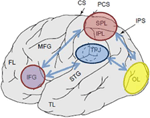
Figura 1. Reprezentarea schematică a reperelor anatomice relevante proiectate pe suprafața laterală a creierului uman. Lobul parietal Superior (SPL) și lobul parietal inferior (IPL) sunt regiuni din partea dorsală a cortexului parietal care subservează orientarea atențională de sus în jos. Joncțiunea Temporo-parietală (TPJ) este o regiune din cortexul parietal ventral care subservează orientarea atențională de jos în sus. Se crede că ambele, SPL și TPJ, provoacă semnale de control responsabile pentru modulațiile atenționale ulterioare observate asupra regiunilor senzoriale, în acest caz modulând (etichetate cu săgeți Albastru închis) activitate evocată vizual în lobul occipital (OL). În plus, se consideră că zonele de-a lungul girusului frontal inferior (IFG) și joncțiunii frontale inferioare (IFJ) servesc drept zone de convergență pentru controlul atențional condus de stimul și de sus în jos (marcate de săgeți bidirecționale albastre deschise).
într-o sarcină tipică menită să angajeze alocarea atențională de sus în jos, indivizilor li se arată două fluxuri de prezentare vizuală serială rapidă (RSVP) poziționate periferic și sunt instruiți inițial să monitorizeze un flux pentru un indiciu (de exemplu, o cifră între fluxul de Litere). Identitatea indicelui indică dacă subiectul trebuie să mențină atenția asupra fluxului curent sau să transfere atenția către celălalt flux (Yantis și colab., 2002; Yantis și Serences, 2003). Două descoperiri majore sunt observate în astfel de paradigme. Primul are legătură cu activarea crescută în regiunile senzoriale reprezentând locația participată la moment (de exemplu, activitate crescută în regiunile vizuale primare stângi atunci când este participat fluxul RSVP drept). Această constatare oferă dovezi ferme că participanții participă la o anumită locație și că atenția modulează puterea răspunsului senzorial (vezi Figura 1; Moran și Desimone, 1985; O ‘ Craven și colab., 1997). A doua constatare are legătură cu observația că regiunile dorsale ale lobului parietal sunt activate selectiv prin schimbări de atenție de sus în jos. Se observă că timpul de activitate SPL/IPL este de natură tranzitorie, sugerând că această zonă a cortexului parietal este sursa unui scurt semnal de control atențional pentru a schimba stările atente într-o manieră de sus în jos (Yantis și colab., 2002).
Mai multe studii fMRI au documentat că captarea atențională de jos în sus, mediată de proeminența stimulului și/sau relevanță, este subservată de joncțiunea temporo-parietală (TPJ; Figura 1). De exemplu, atunci când subiecții participă și monitorizează o modificare a stimulului vizual sau auditiv, prezentată simultan, activarea regiunilor TPJ ale lobului parietal este îmbunătățită. Pe lângă sensibilitatea aparentă la stimulii relevanți, TPJ este activat și ca răspuns la evenimente potențial noi (neașteptate sau rare) atunci când un organism este angajat într-un context comportamental neutru sau când este angajat într-o sarcină (Marois și colab., 2000; Downar și colab., 2002; Serences și colab., 2005; Corbetta și colab., 2008; Asplund și colab., 2010; Diquattro și Geng, 2011; Geng și Mangun, 2011). Această activare are loc independent de modalitatea (auditivă, tactilă și vizuală) în care este livrată intrarea, reflectând natura multisenzorială a TPJ (dar vezi Downar și colab., 2001).
într-o sarcină tipică care examinează mecanismul neuronal al captării atenționale de jos în sus, participanților li se prezintă un flux RSVP de elemente în centrul afișajului și li se cere să identifice o țintă predefinită (de exemplu, să identifice litera roșie prezentată într-un flux RSVP de non-ținte albe). O anumită proporție de studii conține un distractor important irelevant pentru sarcină prezentat la diferite intervale de timp înainte de debutul țintei, în timp ce alte studii conțin doar distractorul important (adică fără țintă). Studiile „țintă-distractor” sunt utilizate pentru a testa gradul de captare, arătând că distractorul irelevant pentru sarcină este de fapt important, rezultând astfel o scădere a preciziei țintei. Studiile „distractor-in-isolation” sunt utilizate pentru analize suplimentare, deoarece astfel de studii permit examinarea activității provocate distractorului important, fără contaminare din procesele legate de țintă. Câteva descoperiri importante apar din astfel de paradigme. În primul rând, atunci când distractorii sunt separați spațial de locația țintă, distractorii de captare sunt însoțiți de o activitate corticală crescută în regiunile corespunzătoare ale cortexului senzorial (de exemplu, cortexul vizual organizat retinotopic; vezi Figura 1). Astfel de rezultate oferă dovezi puternice că, în timpul captării, atenția spațială este de fapt captată la locația spațială ocupată de distractor (Serences și colab., 2005). În al doilea rând, regiunile ventrale ale cortexului parietal, în principal în cadrul TPJ, sunt activate selectiv prin schimbări de atenție de jos în sus, involuntare. La fel ca activitatea din SPL pentru orientarea de sus în jos, cursul de timp al activității observat peste TPJ este de natură tranzitorie, sugerând că această regiune este sursa unui scurt semnal de control atențional pentru a schimba atenția într-o manieră de jos în sus.trebuie remarcat faptul că, în timp ce această revizuire se concentrează pe abordarea funcțiilor cognitive ale cortexului parietal posterior, alte regiuni, în special cele din cortexul frontal, sunt, de asemenea, recrutate pentru alocarea atențională de sus în jos și de jos în sus. Astfel de regiuni includ cortexul frontal ventral (VFC), câmpurile oculare frontale (FEF), joncțiunea frontală inferioară (IFJ) și girusul frontal inferior (IFG; Corbetta și Shulman, 2002, 2011; Serences și colab., 2005; Asplund și colab., 2010; Diquattro și Geng, 2011).
rolul lobului Parietal în selecția de sus în jos și de Jos în sus: Neuropsihologie
istoric cercetătorii s-au bazat critic pe studiile neuropsihologice ale pacienților cu neglijare hemispațială (o tulburare de alocare spațială a atenției la hemi-spațiul stâng) pentru a obține o perspectivă asupra funcțiilor cognitive asociate cu lobul parietal. În literatura neuropsihologică clasică, cortexul parietal, ca întreg, a fost în general considerat locul leziunii primare pentru neglijarea hemispațială. Această viziune, elaborată în detaliu de primii cercetători (Critchley, 1953; McFire și Zangwill, 1960; Piercy, 1964) a recunoscut în mod clar asocierea dintre leziunea parietală și neglijarea care a urmat. Această perspectivă a avut loc în mare parte prin anii 1980, când Posner și colegii (1984) au folosit paradigma ascunsă vizuo-spațială pentru a arăta că deteriorarea lobului parietal produce un deficit în operația de „dezactivare” (retragerea atenției dintr-o locație și mutarea acesteia în alta) atunci când ținta este contralaterală leziunii. Cu toate acestea, în ciuda acestui progres major în înțelegerea bazei neuronale a atenției și în special a rolului de „decuplare” al cortexului parietal, descoperirile lor presupun un singur sit cortical (cortexul parietal) și o singură capacitate funcțională („decuplare”). Spre deosebire de această abordare mai monolitică a creierului (cortexul parietal) și a comportamentului (dezangajarea atențională), lucrările recente de comportament și neuroimagistică (revizuite mai sus și în altă parte) sugerează că atât Regiunea corticală, cât și comportamentul atențional asociat pot fi subdivizate în profiluri calitativ diferite.
având în vedere segregarea rețelelor corticale în procese de sus în jos și de jos în sus, o predicție evidentă este că deteriorarea porțiunilor superioare ale lobului parietal (SPL SUBSUMANT) ar trebui să producă un deficit de orientare atențională direcționată spre obiectiv, în timp ce deteriorarea porțiunilor inferioare ale lobului parietal (TPJ subsumant) ar duce la un deficit asociat cu captarea atenției stimulate. În măsura în care aceste corespondențe creier-comportament au fost explorate în literatura neuropsihologică, această predicție nu este în mod evident susținută. De exemplu, simptomele clinice ale neglijării hemispațiale sunt puternic asociate cu deteriorarea porțiunilor inferioare ale lobului parietal, care include TPJ, mai degrabă decât cu porțiuni superioare precum SPL (Friedrich și colab., 1998; Shomstein și colab., 2010; Corbetta și Shulman, 2011). Acest lucru este oarecum în contradicție cu literatura de neuroimagistică, ceea ce sugerează că rolul TPJ este în captarea atenției, mai degrabă decât în orientarea voluntară a atenției, domeniul în care pacienții neglijați par să aibă cea mai mare dificultate. Pentru a complica lucrurile în continuare, s-a observat că leziunile care implică exclusiv SPL, produc rareori dovezi clinice de neglijare (Vallar și Perani, 1986). Un alt studiu recent cu pacienți cu leziuni centrate în principal pe TPJ și STG, dar conservate SPL, Corbetta și colab. (2005) a arătat că neglijarea spațială, precum și recuperarea acesteia, au fost asociate cu restabilirea activității atât în regiunile ventrale temporo-parietale, cât și în cele parietale dorsale (vezi Corbetta și Shulman, 2011 pentru o revizuire). Deși interesant și interesant în concluziile sale, acest ultim studiu nu diferențiază contribuția relativă a căilor dorsale și ventrale la diferite tipuri de atenție, deoarece pacienții au fost testați doar pe o variantă a sarcinii de atenție spațială ascunsă Posner, sarcină despre care se crede că implică atât orientarea atențională de sus în jos, cât și de jos în sus.
pentru a distinge între controlul atențional condus de obiective și captarea atențională importantă și pentru a examina cartografierea lor pe SPL și, respectiv, TPJ, studiul recent a adoptat două paradigme comportamentale, fiecare vizând una dintre aceste forme de atenție (Shomstein și colab., 2010). Pentru a examina integritatea orientării atenționale de sus în jos la pacienți, a fost utilizată o sarcină de sus în jos care impune participanților să transfere atenția spațială între fluxurile RSVP separate spațial (o sarcină care a fost utilizată cu succes pentru a demonstra activarea SPL în studiile fMRI (Yantis și colab., 2002)). În mod similar, pentru a examina abilitățile de orientare atențională de jos în sus ale pacienților, o variantă a Folk și colab. (2002) paradigma de captare contingentă a fost utilizată în care participanții au detectat ținte care au apărut la fixare în timp ce singletonii de culoare irelevanți ai sarcinii au fost străluciți în periferie. Măsura în care distractorii irelevanți ai sarcinii interferează cu sarcina centrală de detectare a fost apoi utilizată ca măsură a captării atenționale de jos în sus (Bacon și Egeth, 1994; Folk și colab., 2002).
predicțiile au fost următoarele: pacienții cu leziuni ale porțiunilor superioare ale lobului parietal (afectând SPL) ar trebui să fie afectați în sarcina de orientare atențională de sus în jos (cu performanțe conservate în sarcina de captare), în timp ce pacienții cu leziuni ale porțiunilor inferioare ale lobului parietal (afectând TPJ) ar trebui să fie afectați în sarcina de captare (cu performanțe reduse în sarcina de sus în jos). O dublă disociere a acestei forme nu numai că atestă componentele independente ale atenției, dar sugerează, de asemenea, că astfel de componente atenționale sunt mediate de mecanisme neuronale independente. Opt pacienți cu neglijare vizuo-spațială au fost recrutați pentru studiu și au finalizat două sarcini, atingând fie orientarea atențională bazată pe stimul, fie orientarea spre obiectiv. Pe baza profilului lor comportamental, pacienții au fost sortați în grupuri și s-a explorat suprapunerea leziunilor (figura 2a). Pacienții care au prezentat dificultăți în orientarea atențională direcționată spre obiectiv, cuantificată prin indicele atențional de sus în jos (figura 2b), au prezentat o suprapunere a leziunii centrată pe porțiuni superioare ale lobului parietal (SPL SUBSUMANT) cu lobul parietal inferior cruțat (TPJ). Pacienții cu leziune se suprapun centrat pe porțiunile inferioare ale lobului parietal (subsumând TPJ), dar cruțați SPL au efectuat în mod normal pe sarcina de orientare orientată spre obiectiv, rămânând în același timp imuni la captarea atențională (figura 2c). Constatările din acest studiu sugerează în mod clar că SPL și TPJ sunt regiuni anatomice care sunt recrutate în mod necesar în scopul orientării de sus în jos și de jos în sus și că deteriorarea SPL și TPJ duce la tulburări de orientare de sus în jos și respectiv de jos în sus.
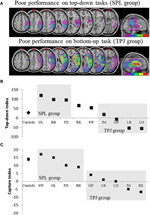
Figura 2. Rezultatele studiului neuropsihologic au vizat investigarea contribuției relative a SPL și TPJ la orientarea de sus în jos și de jos în sus. (A) suprapuneri de leziuni (suprapunere minimă purpurie; suprapunere maximă roșie) pentru pacienții grupați după deficite comportamentale în orientarea atențională de sus în jos, etichetați grupul SPL (panoul superior); și pacienții grupați după deficite comportamentale în orientarea de jos în sus, etichetați grupul TPJ (panoul inferior). (B) performanța comportamentală pe sarcina de sus în jos rezumată cu un „indice de sus în jos” care cuantifică diferențele dintre deplasările spațiale de sus în jos făcute de la stânga la dreapta și invers. Controalele și grupul lezionat TPJ prezintă eficiențe similare în executarea schimbărilor spațiale, în timp ce pacienții cu leziuni SPL prezintă o eficiență scăzută. Controlul grupului și datele individuale ale pacientului (etichetate cu inițialele pacientului) sunt reprezentate pe abscisă. (C) „indicele de captare” este o măsură a atenției de jos în sus și cuantifică măsura în care distractorii irelevanți ai sarcinii captează atenția departe de sarcină. Controalele și grupul lezionat SPL prezintă valori de captare similare, astfel încât ambele grupuri sunt capturate de distractorii irelevanți ai sarcinii. Grupul lezionat TPJ arată un indice de captare mult mai redus (eșecul de a fi capturat). Rețineți că pacienții au fost plasați în grupul SPL sau TPJ pe baza comportamentului, mai degrabă decât pe baza leziunii, notați astfel consistența cu care pacienții ajung în grupul corespunzător.
interacțiunea dintre selecția de sus în jos și de jos în sus
deși se pare că există o asociere puternică între orientarea orientată spre obiective și SPL și orientarea bazată pe stimuli și TPJ, datele de la Shomstein și colab. (2010) studiul pacientului sugerează că aceste două sisteme nu sunt complet independente. Această concluzie este susținută de constatarea că pacienții cu leziuni SPL au prezentat un model de performanță etichetat ca „hiper captare.”Spre deosebire de controale, pentru care doar distractorul colorat țintă a captat atenția (ducând la o precizie mai mică a țintei), distractorii colorați irelevanți s-au dovedit a distrage atenția pentru pacienții cu leziune SPL. În plus, în timp ce pentru controale atenția a fost captată de distractori numai atunci când au precedat debutul țintei, pentru pacienții cu leziuni SPL atenția a fost chiar captată de distractorii prezentați simultan cu ținta. Acest model de performanță poate fi explicat prin următorul cadru: SPL este responsabil pentru orientarea de sus în jos a atenției care include determinarea aspectelor stimulilor care sunt relevante pentru sarcină (de exemplu, căutarea țintei roșii; Corbetta și Shulman, 2002; Serences și colab., 2005). Acest set atențional constrânge apoi TPJ, astfel încât mecanismul de captare a atenției care este mediat de TPJ este declanșat doar de informațiile relevante ale sarcinii (de exemplu, distractorii roșii care captează atenția și distractorii gri care nu captează atenția atunci când caută o țintă roșie). Absența SPL împiedică stabilirea unui set atențional relevant pentru sarcină și, prin urmare, orice stimul, sarcină relevantă sau nu, este considerat important, prin urmare, captarea atenției (de exemplu, distractor irelevant pentru sarcină captarea atenției pentru grupul SPL) fără discriminare.s-a sugerat că SPL și TPJ ar putea interacționa în cel puțin unul din cele două moduri posibile. Prima posibilitate este că TPJ servește ca un sistem de alertare care detectează stimuli relevanți din punct de vedere comportamental, dar nu are rezoluția spațială ridicată, astfel când este detectat un stimul relevant din punct de vedere comportamental, locația sa precisă este furnizată de SPL care stochează hărți spațiale (Kastner și colab., 1999; Wojciulik și Kanwisher, 1999; Bisley și Goldberg, 2003; Silver și colab., 2005). O posibilitate ipotetică legată este că mecanismul de captare (care include TPJ) acționează ca un întrerupător al activității cognitive în curs de desfășurare atunci când este prezentat un stimul relevant din punct de vedere comportamental (Corbetta și Shulman, 2002, 2011). Modelul de activitate” hiper-captare ” observat la pacienții cu TPJ conservat, dar SPL lezat, oferă dovezi suplimentare pentru ipoteza că consideră TPJ ca emitând un semnal de control care termină sarcina la îndemână, servind astfel ca întrerupător de circuit (Corbetta și Shulman, 2002; Serences și colab., 2005). Alte studii recente de neuroimagistică care utilizează diferite paradigme au furnizat dovezi suplimentare pentru o relație interactivă între orientarea atențională de sus în jos și de jos în sus și, ulterior, pentru relația dintre SPL și TPJ (Serences și colab., 2005; Asplund și colab., 2010; Diquattro și Geng, 2011).în timp ce dovezile pentru o interacțiune între cele două sisteme atenționale și cele două substraturi atenționale (SPL și TPJ) sunt puternice, ceea ce rămâne neclar este dacă această interacțiune este directă între SPL și TPJ sau dacă este realizată prin alte regiuni intermediare. Așa cum am menționat mai devreme, rețelele de orientare atențională de sus în jos și de jos în sus angajează diferite regiuni din cortexul frontal, astfel este rezonabil să se presupună că convergența dintre cele două sisteme ar putea fi realizată prin intermediul lobului frontal. Două studii recente care investighează interacțiunea dintre selecția atențională de sus în jos și de jos în sus au furnizat dovezi pentru IFJ și IFG ca posibile site-uri de convergență între selecția bazată pe stimuli și cea direcționată către obiective (Asplund și colab., 2010; Diquattro și Geng, 2011). IFJ și IFG par a fi candidați ideali pentru o astfel de interacțiune, având în vedere implicarea lor generală în atenție și controlul cognitiv, precum și implicarea sa atât în selecția spațială, cât și în cea non-spațială (Koechlin și colab., 2003; Brass și colab., 2005).
rolul lobului Parietal în selecția de sus în jos și de Jos în sus: Fiziologie
în timp ce accentul acestei revizuiri a fost pus în principal pe studiile umane, o mare bogăție de cunoștințe despre implicarea cortexului parietal în orientarea atențională a fost obținută din investigațiile de fiziologie a maimuțelor (vezi revizuirea recentă de Bisley și Goldberg, 2010). Cu toate acestea, atunci când vine vorba de examinarea contribuțiilor relative ale diferitelor regiuni din cortexul parietal la orientarea atențională de sus în jos și de jos în sus, literatura de fiziologie a maimuțelor nu reușește. Motivul principal pentru aceasta este că în cortexul maimuței nu pare să existe dovezi pentru aceeași segregare a controlului de sus în jos și de jos în sus. În schimb, zona intraparietală laterală (buza) considerată inițial a fi implicată în planificarea saccade (Gnadt și Andersen, 1988) este implicată în atenția vizuală și acționează ca o hartă prioritară în care stimulii externi sunt reprezentați în funcție de prioritatea lor comportamentală derivată fie de sus în jos, fie de jos în sus (Colby și Goldberg, 1999; Bisley și Goldberg, 2003, 2010; Balan și Gottlieb, 2006; Ipata și colab., 2006; Buschman și Miller, 2007; Gottlieb și Bălan, 2010).
concluzie
deși se știe mult mai puțin despre cortexul parietal uman decât cel al cortexului omolog de maimuță, studii recente, folosind neuroimagistică și metode neuropsihologice, au început să elucideze distincțiile funcționale și structurale din ce în ce mai fine. Această revizuire s-a concentrat pe studii neuroimagistice și neuropsihologice recente care elucidează rolurile cognitive ale regiunilor dorsale și ventrale ale cortexului parietal în orientarea atențională de sus în jos și de jos în sus și asupra interacțiunii dintre cele două mecanisme de alocare atențională.
Declarație privind conflictul de interese
autorul declară că cercetarea a fost realizată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
mulțumiri
această lucrare a fost susținută de Institutul Național de sănătate Grant EY021644.
Asplund, C. L., Todd, J. J., Snyder, A. P. și Marois, R. (2010). Un rol central pentru cortexul prefrontal lateral în atenția orientată spre obiectiv și stimulată. Nat. Neuroști. 13, 507–512.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Bacon, W. F. și Egeth, H. E. (1994). Captare atențională bazată pe stimuli. Percepție. Psychophys. 55, 485–496.
Pubmed Abstract/Pubmed Full Text
Balan, P. F., și Gottlieb, J. (2006). Integrarea input-ului exogen într-o hartă dinamică evidențiată prin perturbarea atenției. J. Neurosci. 26, 9239–9249.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Behrmann, M., Geng, J. J. și Shomstein, S. (2004). Cortexul Parietal și atenția. Curr. Opin. Neurobiol. 14, 212–217.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Bisley, J. W. și Goldberg, M. E. (2003). Activitatea neuronală în zona intraparietală laterală și atenția spațială. Știință 299, 81-86.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Bisley, J. W. și Goldberg, M. E. (2010). Atenție, intenție și prioritate în lobul parietal. Anu. Rev. Neurologi. 33, 1–21.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Brass, M., Derrfuss, J., Forstmann, B. și von Cramon, D. Y. (2005). Rolul zonei inferioare a joncțiunii frontale în controlul cognitiv. Tendințe Cogn. Sci. 9, 314–316.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Buschman, T. J. și Miller, E. K. (2007). Controlul de sus în jos față de controlul de jos în sus al atenției în cortexurile parietale prefrontale și posterioare. Știință 315, 1860-1862.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Chiu, Y. C. și Yantis, S. (2009). O sursă independentă de domeniu de control cognitiv pentru seturi de sarcini: schimbarea atenției spațiale și schimbarea regulilor de clasificare. J. Neurosci. 29, 3930–3938.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Colby, C. L. și Goldberg, M. E. (1999). Spațiu și atenție în cortexul parietal. Anu. Rev. Neurologi. 22, 319–349.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., Kincade, J. M., Ollinger, J. M., McAvoy, mp, și Shulman, G. L. (2000). Orientarea voluntară este disociată de detectarea țintei în cortexul parietal posterior uman. Nat. Neuroști. 3, 292–297.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., Kincade, M. J., Lewis, C., Snyder, A. Z. și Sapir, A. (2005). Baza neuronală și recuperarea deficitelor de atenție spațială în neglijarea spațială. Nat. Neuroști. 8, 1603–1610.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M., Patel, G. și Shulman, G. L. (2008). Sistemul de reorientare a creierului uman: de la mediu la teoria minții. Neuronul 58, 306-324.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M. și Shulman, G. L. (2002). Controlul atenției orientate spre obiective și stimulate în creier. Nat. Rev. Neurologi. 3, 201–215.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Corbetta, M. și Shulman, G. L. (2011). Neglijarea spațială și rețelele de atenție. Anu. Rev. Neurologi. 34, 569–599.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Critchley, M. (1953). Lobii Parietali. Londra, Marea Britanie: Hafner Press.
Pubmed Abstract/Pubmed Full Text
Diquattro, N. E. și Geng, J. J. (2011). Cunoașterea contextuală configurează rețelele de control atențional. J. Neurosci. 31, 18026–18035.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Downar, J., Crawley, A. P., Mikulis, DJ și Davis, K. D. (2001). Efectul relevanței sarcinii asupra răspunsului cortical la modificările stimulilor vizuali și auditivi: un studiu fMRI legat de evenimente. Neuroimagine 14, 1256-1267.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Downar, J., Crawley, A. P., Mikulis, DJ și Davis, K. D. (2002). O rețea corticală sensibilă la saliența stimulului într-un context comportamental neutru în mai multe modalități senzoriale. J. Neurofiziol. 87, 615–620.
Pubmed Abstract/Pubmed Full Text
Drummond, L. și Shomstein, S. (2010). Atenție bazată pe obiecte: schimbare sau incertitudine? Atten. Percepție. Psychophys. 72, 1743–1755.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Egeth, H. E. și Yantis, S. (1997). Atenție vizuală: control, reprezentare și curs de timp. Anu. Rev. Psychol. 48, 269–297.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Eriksen, C. W. și Hoffman, J. E. (1972). Caracteristicile temporale și spațiale ale codificării selective din afișajele vizuale. Percepție. Psychophys. 12, 201–204.
Folk, C. L., Leber, A. B. și Egeth, H. E. (2002). Te-a făcut să clipești! Captarea atențională contingentă produce o clipire spațială. Percepție. Psychophys. 64, 741–753.
Pubmed Abstract/Pubmed Full Text
Folk, C. L., Remington, R. W. și Johnston, J. C. (1992). Orientarea ascunsă involuntară depinde de setările de control atențional. J. Exp. Psihol. Zumzet. Percepție. Efectuați. 18, 1030–1044.
Pubmed Abstract/Pubmed Full Text
Friedrich, F. J., Egly, R., Rafal, R. D. și Beck, D. (1998). Deficite de atenție spațială la om: o comparație a leziunilor superioare ale joncțiunii parietale și temporo-parietale. Neuropsihologie 12, 193-207.
Pubmed Abstract/Pubmed Full Text
Geng, J. J. și Behrmann, M. (2002). Probabilitatea localizării țintă facilitează căutarea vizuală implicit la participanții normali și la pacienții cu neglijare hemispațială. Psihol. sci. 13, 520–525.
Pubmed Abstract/Pubmed Full Text
Geng, J. J. și Behrmann, M. (2005). Probabilitatea spațială ca un indiciu atențional în căutarea vizuală. Percepție. Psychophys. 67, 1252–1268.
Pubmed Abstract/Pubmed Full Text
Geng, J. J. și Mangun, G. R. (2011). Activarea joncțiunii temporoparietale drepte printr-un indiciu contextual important facilitează discriminarea țintă. Neuroimagine 54, 594-601.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Gibson, B. S. și Kelsey, E. M. (1998). Captarea atențională bazată pe stimul este condiționată de setul atențional pentru caracteristicile vizuale displaywide. J. Exp. Psihol. Zumzet. Percepție. Efectuați. 24, 699–706.
Pubmed Abstract/Pubmed Full Text
Giesbrecht, B., Woldorff, M. G., Song, A. W. și Mangun, G. R. (2003). Mecanisme neuronale de control de sus în jos în timpul atenției spațiale și caracteristice. Neuroimagine 19, 496-512.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Gnadt, J. W. și Andersen, R. A. (1988). Activitatea de planificare motorie legată de memorie în cortexul parietal posterior al macacului. Exp. Brain Res. 70, 216-220.
Pubmed Abstract/Pubmed Full Text
Gottlieb, J. și Balan, P. (2010). Atenția ca decizie în spațiul informațional. Tendințe Cogn. Sci. 14, 240–248.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Greenberg, A. S., Esterman, M., Wilson, D., Serences, J. T. și Yantis, S. (2010). Controlul atenției spațiale și bazate pe caracteristici în cortexul frontoparietal. J. Neurosci. 30, 14330–14339.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Hopfinger, J. B., Buonocore, M. H. și Mangun, G. R. (2000). Mecanismele neuronale ale controlului atențional de sus în jos. Nat. Neuroști. 3, 284–291.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Ipata, A. E., Gee, A. L., Gottlieb, J., Bisley, J. W. și Goldberg, M. E. (2006). Răspunsurile buzelor la un stimul popout sunt reduse dacă este ignorat în mod deschis. Nat. Neuroști. 9, 1071–1076.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Johnston, W. A. și Dark, V. J. (1986). Atenție Selectivă. Anu. Rev. Psychol. 37, 43–75.
Juola, J. F., Koshino, H. și Warner, C. B. (1995). Compromisuri între efectele atenționale ale indiciilor spațiale și onsets abrupte. Percepție. Psychophys. 57, 333–342.
Pubmed Abstract/Pubmed Full Text
Kastner, S., Pinsk, M. A., de Weerd, P., Desimone, R. și Ungerleider, L. G. (1999). Activitate crescută în cortexul vizual uman în timpul atenției direcționate în absența stimulării vizuale. Neuron 22, 751-761.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Koechlin, E., Ody, C. și Kouneiher, F. (2003). Arhitectura controlului cognitiv în cortexul prefrontal uman. Știință 302, 1181-1185.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Koshino, H., Warner, C. B. și Juola, J. F. (1992). Eficacitatea relativă a indiciilor centrale, periferice și cu debut brusc în atenția vizuală. Q. J. Exp. Psihol. Un 45, 609-631.
Pubmed rezumat/Pubmed textul integral
Liu, T., Slotnick, S. D., Serences, J. T., și Yantis, S. (2003). Mecanisme corticale de control atențional bazat pe caracteristici. Cereb. Cortex 13, 1334-1343.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Marois, R., Leung, H. C. și Gore, J. C. (2000). O abordare bazată pe stimuli pentru identitatea obiectului și procesarea locației în creierul uman. Neuron 25, 717-728.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
McFire, J. și Zangwill, O. L. (1960). Dizabilități Visuo-constructive asociate cu leziuni ale emisferei cerebrale stângi. Creierul 82, 243-260.
Moore, C. M. și Egeth, H. (1998). Cum afectează atenția bazată pe caracteristici procesarea vizuală? J. Exp. Psihol. Zumzet. Percepție. Efectuați. 24, 1296–1310.
Pubmed Abstract/Pubmed Full Text
Moran, J., și Desimone, R. (1985). Porti de atentie selectiva procesare vizuala in cortexul extrastriat. Știință 229, 782-784.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
O ‘ Craven, K., Rosen, B. R., Kwong, K. K., Treisman, A. și Savoy, R. L. (1997). Atenția voluntară modulează activitatea fMRI în Mt-MST umană. Neuron 18, 591-598.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Piercy, M. (1964). Efectele leziunilor cerebrale asupra funcției intelectuale: o revizuire a tendințelor actuale de cercetare. Br. J. Psihiatrie 110, 310-352.
Pubmed Abstract/Pubmed Full Text
Posner, M. I. (1980). Orientarea atenției. Q. J. Exp. Psihol. 32, 3–25.
Pubmed Abstract/Pubmed Full Text
Posner, M. I., Snyder, C. R. și Davidson, B. J. (1980). Atenție și detectarea semnalelor. J. Exp. Psihol. 109, 160–174.
Pubmed Abstract/Pubmed Full Text
Posner, M. I., Walker, J. A., Friedrich, F. J. și Rafal, R. D. (1984). Efectele leziunilor parietale asupra orientării ascunse a atenției. J. Neurosci. 4, 1863–1874.
Pubmed Abstract/Pubmed Full Text
Serences, J. T., Shomstein, S., Leber, A. B., Golay, X., Egeth, H. E. și Yantis, S. (2005). Coordonarea controlului atențional voluntar și stimulat în cortexul uman. Psihol. Sci. 16, 114–122.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Shaw, M. L. (1978). Un model de alocare a capacității pentru timpul de reacție. J. Exp. Psihol. Zumzet. Percepție. Efectuați. 4, 586–598.
Shomstein, S. și Behrmann, M. (2006). Sisteme corticale care mediază atenția vizuală atât asupra obiectelor, cât și asupra locațiilor spațiale. Proc. Natl. Acad. Sci. SUA 103, 11387-11392.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Shomstein, S., Lee, J. și Behrmann, M. (2010). Orientare atențională de sus în jos și de jos în sus: investigarea rolului cortexului parietal dorsal și ventral. Exp. Brain Res. 206, 197-208.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Shomstein, S. și Yantis, S. (2004a). Prioritizarea configurală și contextuală în atenția bazată pe obiecte. Psychon. Taur. 11, 247-253.
Pubmed Abstract/Pubmed Full Text
Shomstein, S. și Yantis, S. (2004b). Controlul atenției se schimbă între viziune și audiție în cortexul uman. J. Neurosci. 24, 10702–10706.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Shomstein, S. și Yantis, S. (2006). Cortexul Parietal mediază controlul voluntar al atenției auditive spațiale și nonspațiale. J. Neurosci. 26, 435–439.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Silver, M. A., Ress, D., și Heeger, D. J. (2005). Hărți topografice ale atenției spațiale vizuale în cortexul parietal uman. J. Neurofiziol. 94, 1358–1371.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Tamber-Rosenau, B. J., Esterman, M., Chiu, Y.-C. și Yantis, S. (2011). Mecanisme corticale de control cognitiv pentru schimbarea atenției în viziune și memoria de lucru. J. Cogn. Neuroști. 23, 2905–2919.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Theeuwes, J. (1991). Controlul exogen și endogen al atenției: efectul de onsets vizuale și compensări. Percepție. Psychophys. 49, 83–90.
Pubmed Abstract/Pubmed Full Text
Vallar, G. și Perani, D. (1986). Anatomia neglijării unilaterale după leziunile accident vascular cerebral din emisfera dreaptă. Un studiu de corelație clinic/CT-scan la om. Neuropsihologia 24, 609-622.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Wojciulik, E. și Kanwisher, N. (1999). Generalitatea implicării parietale în atenția vizuală. Neuronul 23, 747-764.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Yantis, S. și Jonides, J. (1984). Întreruperi vizuale abrupte și atenție selectivă: dovezi din căutarea vizuală. J. Exp. Psihol. Zumzet. Percepție. Efectuați. 10, 601–621.
Pubmed Abstract/Pubmed Full Text
Yantis, S. și Jonides, J. (1990). Întreruperi vizuale abrupte și atenție selectivă: alocare voluntară versus automată. J. Exp. Psihol. Zumzet. Percepție. Efectuați. 16, 121–134.
Pubmed Abstract/Pubmed Full Text
Yantis, S., Schwarzbach, J., Serences, J. T., Carlson, R. L., Steinmetz, M. A., Pekar, J. J. și Courtney, S. M. (2002). Activitate neuronală tranzitorie în cortexul parietal uman în timpul schimbărilor de atenție spațială. Nat. Neuroști. 5, 995–1002.
Pubmed Abstract | Pubmed Full Text/CrossRef Full Text
Yantis, S. și Serențe, J. T. (2003). Mecanisme corticale de control atențional bazat pe spațiu și pe obiecte. Curr. Opin. Neurobiol. 13, 187–193.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text