Introducere
resursele fosile sunt principala materie primă pentru energie și compuși organici, iar utilizarea lor are ca rezultat emisia de gaze cu efect de seră asociate schimbărilor climatice. Următorul accident climatic necesită o tranziție urgentă de la o economie bazată pe fosile la o economie bazată pe bio în care biomasa lignocelulozică, mai degrabă decât petrolul, este utilizată pentru producția de combustibili, substanțe chimice și materiale. Lemnul este o sursă importantă de biomasă lignocelulozică; este compus în principal din pereți celulari secundari îngroșați, bogați în celuloză, hemiceluloze și lignină. Toți cei trei polimeri pot fi valorificați în economia bazată pe bio. Celuloza este o sursă pentru industria celulozei și hârtiei și atât celuloza, cât și hemicelulozele pot fi depolimerizate în monozaharidele lor pentru fermentare, de exemplu, bioetanol, acid lactic și detergenți (Vanholme și colab., 2013b). Deoarece lignina afectează negativ eficiența prelucrării lemnului în aceste aplicații, copacii pot fi proiectați pentru a acumula mai puțină lignină, pentru a deveni mai susceptibili pentru producerea hârtiei și a zaharurilor fermentabile. Pe de altă parte, lignina este considerată din ce în ce mai mult o componentă valoroasă în economia bazată pe bio. Într-adevăr, având în vedere că lignina este cea mai mare sursă aromatică regenerabilă de pe Pământ, viabilitatea economică a unei bio-rafinării poate fi crescută semnificativ dacă lignina este, de asemenea, valorificată și utilizată ca resursă pentru producerea de substanțe chimice (Holladay și colab., 2007; Tuck și colab., 2012; Davis și colab., 2013; Ragauskas și colab., 2014; Li C. și colab., 2015; Van den Bosch și colab., 2015; Rinaldi și colab., 2016; Upton și Kasko, 2016; Schutyser și colab., 2018).
lignina polimerică este compusă din monolignoli produși de căile biosintetice fenilpropanoid și monolignol, printr-o serie de reacții enzimatice începând cu dezaminarea fenilalaninei (Figura 1). Monolignolii sunt sintetizați în citoplasmă și translocați în apoplast, unde sunt dehidrogenați la radicalii monolignol prin acțiunea lacazelor și peroxidazelor (Berthet și colab., 2011; Zhao și colab., 2013). Acești radicali monolignol se cuplează apoi unul cu celălalt într-un mod combinatorial, generând o serie de legături chimice, cum ar fi legătura aril-eter (XV-o-4), legătura rezinol (VIII-XV) și legătura fenilcumarană (5) (Boerjan și colab., 2003; Ralph și colab., 2004; Vanholme și colab., 2010). Cele mai frecvente monolignoli sunt alcooli hidroxicinamilici p-cumaril, coniferil, și alcooli sinapilici, care generează unitățile H, G și s la încorporarea lor în polimer lignină, respectiv (Bonawitz și Chapple, 2010; Ralph și colab., 2019; Vanholme și colab., 2019). Contribuția relativă a blocurilor de construcție a ligninei variază între taxoni, stadiul de dezvoltare, tipul de țesut și celulă și chiar stratul peretelui celular; lignina din rasinoase (gimnosperme) este compusă aproape în întregime din unități G cu o fracțiune minoră de unități H, în timp ce lignina din lemn de esență tare (angiosperme) are unități S în plus față de unitățile G și urme de unități H (Boerjan și colab., 2003; Vanholme și colab., 2010, 2019). Pe lângă acești monolignoli tradiționali, o varietate de alte molecule aromatice p-hidroxilate pot fi încorporate în polimerul ligninei la diferite niveluri (Vanholme și colab., 2019).
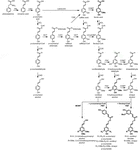
Figure 1. Lignin biosynthetic pathway. Alternative monomers and heterologously expressed enzymes are shown in bold. PAL, phenylalanine ammonia-lyase; C4H, cinnamate 4-hydroxylase; 4CL, 4-coumarate:CoA ligase; HCT, p-Hydroxycinnamoyl-CoA:quinate/shikimate-p-hydroxycinnamoyltransferase; C3’H, p-coumaroyl quinate/shikimate 3’-hydroxylase; CSE, caffeoyl shikimate esterase; CCoAOMT, caffeoyl-CoA O-methyltransferase; CCR, cinnamoyl-CoA reductase; F5H, ferulate 5-hydroxylase/CAld5H, coniferaldehyde 5-hydroxylase; COMT, acid cafeic o-metiltransferază; CAD, alcool cinamilic dehidrogenază; FMT, feruloil-CoA monolignol transferază; PMT, p-cumaroil-CoA monolignol transferază.
având în vedere că lignina este un factor major de recalcitranță în procesele de delignificare a lemnului, s-au depus eforturi mari de cercetare pentru a descoperi calea biosintetică a ligninei și pentru a studia efectele perturbațiilor genelor de biosinteză a ligninei asupra cantității și compoziției ligninei și asupra eficienței procesării lemnului. În timp ce modificările genelor variind de la acei factori de transcripție care codifică până la cei care codifică enzimele oxidative au dus la modificarea conținutului, compoziției sau depunerii ligninei (Eriksson și colab., 2000; Li Y. H. și colab., 2003; Liang și colab., 2008; Lu și colab., 2013; Lin și colab., 2016; Xu și colab., 2017; Yang și colab., 2017; Obudulu și colab., 2018), această revizuire se va concentra pe rezultatele obținute prin ingineria genelor biosintetice ale ligninei.
Ingineria căii ligninei
Tabelul 1 oferă o imagine de ansamblu a diferitelor studii privind genele biosintetice ale ligninei reglate sau mutante în plop, pin, eucalipt și mesteacăn, cu efectele rezultate asupra eficienței procesării lemnului, atunci când sunt determinate. Reducerea activității oricărei etape a căii biosintetice a ligninei, începând de la PAL până la CAD, poate duce la o reducere a conținutului de lignină (Tabelul 1). Mai mulți parametri influențează gradul de reducere a ligninei, cum ar fi gena țintă și gradul de reglare descendentă a activității enzimei, care la rândul său depinde de eficiența construcției de tăcere utilizate, de dimensiunea familiei de gene și de redundanța din cadrul familiei de gene. În general, reglarea descendentă a pașilor de la C4H până la CCR are ca rezultat o reducere mai dramatică a cantității de lignină (Hu și colab., 1999; Meyermans și colab., 2000; Zhong și colab., 2000; Li L. și colab., 2003; Jia și colab., 2004; Lu și colab., 2004; Lepro-Lepro-Lepro și colab., 2007; Coleman și colab., 2008a, b; Bjurhager și colab., 2010; Mansfield și colab., 2012; Ralph și colab., 2012; Min și colab., 2014; Van Acker și colab., 2014; Zhou și colab., 2015, 2018; Saleme și colab., 2017; Xiang și colab., 2017) decât downregulation de F5H, COMT și CAD (Van Doorsselaere și colab., 1995; Baucher și colab., 1996; Lapierre și colab., 1999; Jouanin și colab., 2000; Van Acker și colab., 2014; Wang și colab., 2018). Reducerea ligninei poate fi asociată cu o creștere a S/G, cum ar fi în C3 ‘ h- (Coleman și colab., 2008a; Ralph și colab., 2012) și ccoaomt-copaci reglați în jos (Meyermans și colab., 2000) sau o scădere a raportului S/G, cum ar fi în CSE- (Saleme și colab., 2017) și copaci COMT-downregulated (Van Doorsselaere și colab., 1995; Lapierre și colab., 1999; Jouanin și colab., 2000). Interesant, s-a constatat că plopii cu nivel scăzut de lignină 4CL au o creștere a S/G (Min și colab., 2014; Xiang și colab., 2017), o scădere a S/G (Voelker și colab., 2010; Zhou și colab., 2015) sau rapoarte comparabile cu tipul sălbatic (Hu și colab., 1999; Li L. și colab., 2003). Această varianță nu poate fi asociată cu promotorul sau cu metoda utilizată pentru reglarea descendentă, sugerând că diferențele în gradul de tăcere, condițiile de creștere sau starea de dezvoltare influențează această trăsătură. Pe de altă parte, cele mai puternice efecte asupra compoziției de lignină H/G/s au fost observate pentru copacii reglați în jos în C3 ‘ H și HCT, care depozitează lignină cu creșteri mari ale conținutului de unitate H (Coleman și colab., 2008a; Ralph și colab., 2012; Vanholme și colab., 2013a), în timp ce copacii care supraexprimă f5h produc lignină puternic îmbogățită în unitățile S (Franke și colab., 2000; Li L. și colab., 2003; Stewart și colab., 2009), și copaci care sunt downregulated în COMT au redus dramatic biosinteza unității S (Van Doorsselaere și colab., 1995; Lapierre și colab., 1999).

Tabelul 1. Prezentare generală a copacilor forestieri cu expresie modificată a genelor de biosinteză a ligninei.
eficiența procesării biomasei poate fi modificată și prin încorporarea crescută a moleculelor care reprezintă în general componente minore în lignina plantelor de tip sălbatic. Încorporarea acidului ferulic în arborii cu deficit de CCR are ca rezultat formarea legăturilor acetale în polimerul de lignină, care se scindează ușor în pretratările de biomasă acidă (LEPL., 2007; Ralph și colab., 2008; Van Acker și colab., 2014). Într-adevăr, nivelurile de acid ferulic din lignină s-au corelat pozitiv cu o eficiență mai mare de zaharificare (Van Acker și colab., 2014). Încorporarea alcoolului 5-hidroxiconiferilic și a 5-hidroxiconiferaldehidei în lignina Plopilor cu deficit de COMT (Van Doorsselaere și colab., 1995; Lapierre și colab., 1999; Jouanin și colab., 2000; Morreel și colab., 2004; Lu și colab., 2010) dă naștere la legături de benzodioxan, prevenind potențial legăturile covalente dintre lignină și grupările hidroxil polizaharidice (Weng și colab., 2010; Vanholme și colab., 2012; Nishimura și colab., 2018). Pe de altă parte, deficitul de COMT are ca rezultat și o lignină mai condensată datorită nivelurilor relativ ridicate ale legăturilor condensate de la SEC-5 la SEC-5, precum și nivelurilor mai scăzute ale legăturilor de la SEC-o-4, atunci când frecvența unitară S scade. Pulpa chimică a lemnului derivat din plopi puternic reglat în jos pentru COMT a dus la un randament mai mare al pulpei, contrabalansat de conținutul rezidual de lignină din pulpă. Acești copaci aveau o lignină mai mică și un conținut mai mare de celuloză (Jouanin și colab., 2000). Pe de altă parte, plopii care au fost reglați modest pentru COMT au avut o scădere mare a producției de celuloză, probabil pentru că conținutul de lignină a rămas normal, în timp ce lignina a avut o frecvență mai mare a legăturilor condensate care au afectat negativ extracția ligninei (Lapierre și colab., 1999; Pilat și colab., 2002). Încorporarea cinamaldehidelor în polimerul de lignină în arborii cu deficit de CAD are ca rezultat lanțuri de polimeri de lignină mai scurte, deci o proporție mai mare de grupări finale fenolice libere care cresc solubilitatea polimerului în alcali. Încorporarea cinamaldehidelor în polimerul ligninei reduce probabil și interacțiunea covalentă a lanțului alifatic cu hemiceluloza, făcând din nou lignina mai solubilă. În plus, datorită sistemului conjugat extins care este generat atunci când o cinamaldehidă-o-4 se cuplează cu un alt monomer, legătura eterică aromatică a cinamaldehidei încorporate devine mai susceptibilă la scindarea alcalină (Lapierre și colab., 1989; Van Acker și colab., 2017).
polimerizarea ligninei este un proces de cuplare radicală combinatorie, permițând o gamă largă de compuși fenolici să fie încorporați în mod natural în polimerul ligninei (Boerjan și colab., 2003; Vanholme și colab., 2019). Cercetătorii au încercat să adapteze cantitatea și compoziția ligninei pentru a îmbunătăți procesarea biomasei prin exprimarea genelor heterologe, urmărind biosinteza și încorporarea diferiților compuși fenolici compatibili ca monolignoli alternativi în polimerul ligninei (Ralph, 2006; Vanholme și colab., 2012; Mottiar și colab., 2016; Mahon și Mansfield, 2018). Un exemplu este introducerea genelor care codifică enzimele care sunt necesare pentru biosinteza unității S în pin; expresia simultană a f5h, COMT și CAD a introdus cu succes unitățile S în Pinus radiata (Wagner și colab., 2015; Edmunds și colab., 2017). Introducerea genei care codifică un monolignol 4-o-metiltransferază (MOMT4) în plop duce la formarea coniferililor 4-o-metilați și a alcoolilor sinapilici, care nu pot fi încorporați în polimerul ligninei în creștere, deoarece le lipsește gruparea hidroxil aromatică. Acest lucru duce la oprirea polimerizării ligninei și are ca rezultat copaci cu conținut mai mic de lignină și eficiență mai mare de zaharificare (Bhuiya și Liu, 2010; Cai și colab., 2016). Plopii au fost, de asemenea, proiectați pentru a conține legături esterice în coloana vertebrală a polimerului de lignină. Esterii coniferil ferulate au fost introduși în polimer prin expresia unei gene FERULOIL-CoA:MONOLIGNOL transferază (FMT) derivată din Angelica sinensis (Wilkerson și colab., 2014), conducând la o eficiență îmbunătățită a zaharificării în diferite condiții de pretratare (Wilkerson și colab., 2014; Kim și colab., 2017; Bhalla și colab., 2018) și o eficiență îmbunătățită a pulpării kraft în comparație cu tipul sălbatic (Zhou și colab., 2017). Esterii monolignol p-cumarat au fost, de asemenea, proiectați în plop, prin expresia unei gene p-CUMAROIL-CoA de orez:monolignol transferază (PMT), rezultând o frecvență mai mare a legăturilor interunitare rezistente și o frecvență mai mare a unităților terminale G și s cu grupări fenolice libere (Smith și colab., 2015; Sibout și colab., 2016). În timp ce în Arabidopsis expresia heterologă a PMT a dus la o cantitate redusă de lignină însoțită de o eficiență crescută de zaharificare (Sibout și colab., 2016), nu a existat o scădere a cantității de lignină în plop și eficiența zaharificării nu a fost determinată (Smith și colab., 2015).
în timp ce s-a demonstrat că mai multe modificări ale cantității și compoziției ligninei oferă îmbunătățiri în procesarea biomasei, aceste modificări au fost adesea însoțite de o penalizare a randamentului biomasei (LEPL la sută și colab., 2007; Wadenb Inktck și colab., 2008; Wagner și colab., 2009; Voelker și colab., 2010; Stout și colab., 2014; Van Acker și colab., 2014; Sykes și colab., 2015; Zhou și colab., 2018). O metastudie recentă a perturbat 21 de gene de biosinteză a ligninei în P. trichocarpa și a integrat în mod cuprinzător rezultatele datelor transcriptomice, proteomice, fluxomice și fenomice ale 221 de linii. Autorii au concluzionat că creșterea copacilor nu este asociată cu cantitatea de lignină, compoziția subunității sau legăturile specifice (Wang și colab., 2018), ci mai degrabă corelată cu prezența vaselor de xilem prăbușite (Coleman și colab., 2008a, b; Wagner și colab., 2009; Voelker și colab., 2010; Vargas și colab., 2016; de Meester și colab., 2018), activarea unei căi de integritate a peretelui celular (Bonawitz și colab., 2014) și/sau acumularea de inhibitori chimici (Gallego-Giraldo și colab., 2011; Muro-Villanueva și colab., 2019).
în timp ce s-au depus eforturi substanțiale pentru scăderea conținutului de lignină prin reglarea descendentă a genelor biosintetice ale ligninei, studiile privind reglarea ascendentă a căii ligninei și supraproducția ligninei au fost rare. Într-adevăr, rapoartele privind supraexprimarea F5H arată o neschimbată sau chiar o scădere a conținutului de lignină (Huntley și colab., 2003; Li L. și colab., 2003; Stewart și colab., 2009; Mansfield și colab., 2012; Edmunds și colab., 2017). Supraexprimarea CAD și COMT a dus la reducerea la tăcere a genelor, mai degrabă decât la reglarea în sus, sau nu a fost detectat niciun efect asupra nivelurilor de Expresie (Baucher și colab., 1996; Lapierre și colab., 1999; Jouanin și colab., 2000; Lepro-Lepro-Lepro-Lepro și colab., 2007; Van Acker și colab., 2014). Supraexprimarea factorilor de transcripție R2R3-MYB PtoMYB92, PtoMYB216 și PtoMYB74 au dus la straturi suplimentare de xilem, pereți celulari xilem mai groși, precum și depunerea ectopică de lignină, iar plantele au acumulat cu 13-50% mai multă lignină (Tian Q. și colab., 2013; Li C. F. și colab., 2015; Li și colab., 2018). Liniile de supraexpresie MYB au reglat constitutiv genele căii de biosinteză a ligninei și, în timp ce plantele supraexprimând MYB92 și MYB74 au avut o penalizare a biomasei, supraexpresia MYB216 a dus la plante cu până la 50% mai multă lignină și fără fenotip de dezvoltare. Deoarece lignina este considerată din ce în ce mai mult o resursă importantă pentru producția durabilă de substanțe chimice (Cao și colab., 2018) ingineria plantelor care produc lignină în exces ar trebui explorată în continuare.
studii de teren
exemplele discutate mai sus arată clar că ingineria ligninei prin reglarea descendentă sau ascendentă a genelor căii fenilpropanoide – sau expresia genelor heterologe – are potențialul de a crește eficiența procesării biomasei lignocelulozice. Din motive practice și de reglementare, majoritatea studiilor raportează datele obținute din analiza copacilor crescuți într-o seră. Cu toate acestea, experimentele cu copaci crescuți într-o seră nu iau în considerare de obicei procesele de dezvoltare, cum ar fi încetarea creșterii și dormitul. În plus, experimentele cu efect de seră nu oferă o perspectivă suficientă asupra interacțiunii plantei proiectate cu factori de mediu, cum ar fi tipul de sol, vântul și agenții patogeni. Înțelegerea acestor interacțiuni este un pas important în traducerea rezultatelor cercetării către aplicații comerciale. Într – adevăr, corpul de lucru produs de studii pentru care a fost acordată permisiunea de a stabili studii pe teren, evidențiază diferențe importante în fenotip între copacii crescuți în seră și în câmp. Tabelul 1 rezumă rapoartele privind studiile pe teren efectuate cu arbori 4CL, CCoAOMT, CCR, COMT și CAD.
confirmând potențialul biomasei lignocelulozice modificate ca substrat pentru aplicații, mai mulți arbori cultivați pe teren cu lignină au arătat îmbunătățiri în prelucrarea lemnului. Plopii reglați în jos pentru CCoAOMT crescuți timp de 5 ani într-un studiu de teren de la Beijing (China), au arătat o eliberare crescută de glucoză și xiloză la zaharificare (Wang și colab., 2012). Plopii reglementați în jos pentru CCR și crescuți într-un proces de teren în Franța, s-au dovedit a fi mai susceptibili la pulpa chimică kraft (LEPL XV și colab., 2007). Două studii suplimentare pe teren efectuate în Franța și Belgia cu plopi reglementați în jos de CCR au dus la o îmbunătățire de până la 160% a producției de etanol în testele simultane de zaharificare și fermentare (SSF); cu toate acestea, plantele au avut o reducere de până la 50% a biomasei (Van Acker și colab., 2014). Studiile de teren cu plop CAD-downreglated au arătat, de asemenea, rezultate promițătoare. Acești copaci au prezentat puțin mai puțin lignină decât tipul sălbatic și s-au dovedit mai susceptibili la delignificarea kraft (Lapierre și colab., 1999). În mod constant, aceleași linii cultivate în studiile pe teren la scară mai mare din Franța și Regatul Unit au arătat o scădere ușoară a cantității de lignină și o îmbunătățire a pulpării kraft considerată relevantă din punct de vedere comercial, deoarece plantele aveau nevoie de 6% mai puțin alcali pentru a obține o delignificare similară cu cea a copacilor de tip sălbatic (Pilat și colab., 2002).cu toate acestea, rapoartele contradictorii privind randamentul biomasei și eficiența procesării în aval sugerează că acești parametri sunt puternic influențați de factorii de mediu. În timp ce un studiu de teren efectuat în China folosind plopi 4CL reglați în jos a constatat că, chiar și cu o scădere cu 28% a conținutului de lignină în comparație cu tipul sălbatic, copacii au avut o înălțime crescută cu aproximativ 8% (Tian X. M. și colab., 2013), în concordanță cu studiile cu efect de seră (Hu și colab., 1999), alte studii pe teren au constatat că plopii 4CL-reglați în jos au scăzut biomasa și au fost uneori chiar pitici (Voelker și colab., 2010; Stout și colab., 2014; Marchin și colab., 2017). Rapoartele diferă, de asemenea, în ceea ce privește eficiența procesării în aval a lemnului derivat din acești plopi crescuți în câmp 4CL. În timp ce o creștere de până la 100% a recuperării zahărului a fost găsită pentru copacii 4cl1-downregulated (35S-driven antisense 4CL construct) crescuți într-un sit montan din Statele Unite (Xiang și colab., 2017), datele obținute din studiile de teren efectuate în Oregon (Statele Unite) au constatat că pt4cl1 antisens condus de promotor 4cl1 plopii tăcuți nu au avut nicio îmbunătățire a eficienței zaharificării în comparație cu tipul sălbatic (Voelker și colab., 2010). De asemenea, un studiu pe termen lung în Wenling (China), a constatat că plopii 4CL-downregulated nu au prezentat o îmbunătățire semnificativă a randamentului zahărului în comparație cu tipul sălbatic (Wang și colab., 2012). În ambele cazuri din urmă, arborii au prezentat scăderi ușoare ale cantității de lignină, care nu s-au tradus într-o eficiență de procesare mai mare, potențial din cauza concentrației mai mari de extracte care ar putea interfera cu activitatea enzimatică (Voelker și colab., 2010).
studiile de teren au arătat că factorii de mediu pot influența lignificarea și pot restabili trăsăturile la nivelurile de tip sălbatic în comparație cu nivelurile atinse atunci când aceleași plante au fost cultivate în seră. În timp ce copacii 4CL-downregulați au scăzut conținutul de lignină atunci când au fost crescuți în condiții de seră, analiza acelorași plopi antisens 4CL, dar crescuți pe câmp, a arătat adesea că conținutul de lignină a fost crescut în comparație cu copacii crescuți în seră și uneori chiar readus la niveluri de tip sălbatic (Stout și colab., 2014; Xiang și colab., 2017). În mod similar, nivelurile de lignină au fost mult mai puțin reduse la plopii cu deficit de CCR atunci când au fost crescuți pe câmp, comparativ cu atunci când au fost crescuți în seră (Van Acker și colab., 2014). Cel puțin pentru plopii cu deficit de CCR, este posibil ca nivelul de lignificare mai ridicat al arborilor crescuți în câmp să se datoreze faptului că probele de lemn au fost prelevate în timpul iernii. Când creșterea copacilor încetează toamna, copacii mai au timp să-și lignifice complet pereții celulari până când arborele intră în repaus, în comparație cu copacii crescuți în seră care dezvoltă continuu xilem nou. Compoziția ligninei s – a dovedit, de asemenea, că diferă între copacii cu lignină joasă crescuți în seră și în câmp. 4CL-plopii reglați în jos crescuți într-un câmp din Carolina de Nord aveau lignină cu un S/G mai mic decât atunci când aceleași linii erau cultivate în seră (Stout și colab., 2014).luate împreună, aceste rezultate arată că datele obținute de la copacii crescuți în seră nu pot fi extrapolate cu ușurință la copacii crescuți în câmp, susținând necesitatea experimentelor de testare pe teren în diferite locații. Unele linii au prezentat o penalizare de randament, făcându-le mai puțin interesante pentru aplicații, subliniind necesitatea unei mai bune înțelegeri a bazei moleculare a penalizării de randament și a dezvoltării de strategii pentru a depăși această problemă.s-a demonstrat că lignina joacă un rol important în rezistența agenților patogeni (Miedes et al., 2014; Zhao și Dixon, 2014) și joacă un rol esențial în a permite plantei să transporte apă. Acest lucru sugerează că modificările ligninei ar putea avea un impact asupra toleranței la stres a plantelor. Deși sunt necesare investigații suplimentare pentru a aborda pe deplin această posibilitate, reglarea descendentă a 4CL, COMT și CAD în plop nu a modificat dramatic performanța de hrănire a erbivorelor care hrănesc frunzele (Tiimonen și colab., 2005; Brodeur-Campbell și colab., 2006; Hjalten și colab., 2013). Efectul reglării descendente a COMT și CAD în plop asupra interacțiunilor plante-insecte a fost, de asemenea, evaluat pe copacii crescuți pe câmp și s-a demonstrat că arborii modificați cu lignină au avut o incidență normală a vizitării și hrănirii insectelor, precum și răspunsuri normale la agenții patogeni microbieni (Pilat și colab., 2002; Halpin și colab., 2007). Aceste rezultate indică faptul că copacii cu lignină modificată nu suferă neapărat mai mult decât plantele sălbatice de dăunători și boli. Cu toate acestea, profilarea microbiomului bacterian endosferic al lemnului recoltat din plopii crescuți în câmp, CCR, a demonstrat schimbări în comunitatea bacteriană, probabil din cauza abundenței modificate a metaboliților fenolici specifici din xilem (Beckers și colab., 2017).
având în vedere rolul ligninei în funcția și structura xilemului, au fost evaluate relațiile de apă ale câtorva plopi modificați cu lignină scăzută. 4CL-plopii reglați în jos s-au dovedit a avea o conductivitate hidraulică redusă, potențial interferând cu creșterea plantelor (Marchin și colab., 2017). Experimentele de stres hidraulic cu plopi reglate în jos pentru CCR, COMT sau CAD au arătat că aceste plante au o rezistență mai mică la cavitație, menținând în același timp conductivitatea hidraulică normală a xilemului și transportul apei (Awad și colab., 2012). Aceste rezultate sugerează că creșterea mutanților cu lignină scăzută ar putea fi influențată de disponibilitatea apei. Ca și pentru orice hibrid nou obținut din reproducerea clasică, sunt necesare teste pe teren pentru a evalua performanța pe teren și toleranța la stres a copacilor proiectați cu lignină.
perspectivele ingineriei ligninei în arborii Forestieri
performanța instalațiilor fabricate cu lignină pare să fie puternic influențată de condițiile de mediu. Cu toate acestea, nu este clar dacă diferențele observate între copacii crescuți în seră și cei crescuți în câmp sau între copacii crescuți în diferite locații de câmp rezultă din diferite niveluri de suprimare a genelor sau din interacțiunea trăsăturii proiectate cu mediul (GxE). Într-adevăr, reglarea descendentă instabilă este un neajuns al tehnicilor de tăcere a genelor care se bazează pe RNAi. Acest lucru este observat prin observarea variației fenotipului de xilem roșu care se observă atunci când anumite gene de biosinteză a ligninei, cum ar fi CAD, COMT sau CCR, sunt reglate în jos. Culoarea roșie a xilemului nu este adesea uniformă pe tot parcursul xilemului, ci apare mai degrabă în patch-uri care reflectă Niveluri variabile de reducere a zgomotului genetic (LEPL XV și colab., 2007; Voelker și colab., 2010; Van Acker și colab., 2014; Figura 2). În plus, utilizarea metodelor de tăcere a genelor poate duce la reducerea concomitentă a membrilor familiei genetice strâns legate – poate în diferite grade – întunecând interpretările și camuflând efectele reglării descendente a genelor individuale.

Figura 2. Reglarea neregulată a genelor de către RNAi. Fenotipul de xilem roșu neuniform observat pe trunchiurile de plopi cu deficit de CCR (dreapta) cultivate într-un studiu de teren din Belgia. Xilemul roșu indică zonele de downregulation CCR. Lemnul din copaci de tip sălbatic este albicios (stânga).
aceste probleme pot fi acum depășite cu ușurință prin utilizarea tehnologiilor de editare a genelor bazate pe CRISPR care permit mutații stabile ale pierderii funcției (knock-out-uri) în genele țintă specifice, permițând disecția funcției genelor individuale în cadrul familiilor. De exemplu, direcționarea membrilor individuali ai familiei genei 4CL în plop a arătat că 4cl1 este legat de lignificare, în timp ce 4cl2 este implicat în producția de proantocianidină (Zhou și colab., 2015). În plus față de alelele knock-out, editarea genică bazată pe CRISPR permite, de asemenea, crearea de noi alele care conferă o reducere parțială a activității enzimei. Aceasta deschide posibilitatea de a regla fin nivelul activității enzimatice reziduale și de a ocoli penalizarea de randament care este adesea observată atunci când cantitatea de lignină scade sub un nivel de prag. O altă cale promițătoare pentru ingineria ligninei în copacii forestieri, posibilă prin ingineria genomului bazată pe CRISPR, este editarea simultană a mai multor gene (stivuirea alelelor) pentru a optimiza eficiența procesării biomasei, așa cum este exemplificat în Arabidopsis, unde stivuirea mutațiilor transaldolazei (tra) și comt, mutațiile c4h și comt sau mutațiile 4CL și comt au dus la îmbunătățiri aditive și sinergice ale eficienței zaharificării (de Vries și colab., 2018). Într-adevăr, o abordare sistemică în P. trichocarpa prezice că reglarea descendentă concomitentă a PAL și CCoAOMT sau PAL, C3 ‘ H și CCOAOMT va îmbunătăți substanțial proprietățile lemnului și eliberarea zahărului (Wang și colab., 2018).
utilizarea editării genomului pe bază de CRISPR în îmbunătățirea arborelui pentru industria celulozei și hârtiei și a bio-rafinăriei, precum și pentru producerea aromelor de platformă din defalcarea hidrogenolitică a ligninei, va fi cea mai valoroasă atunci când această tehnologie este combinată strategic cu alte tehnici de reproducere (Figura 3). Într-adevăr, o mare variație a cantității de lignină și a compoziției S/G există deja în populațiile naturale de arbori forestieri (Studer și colab., 2011). Având în vedere că ambele trăsături afectează eliberarea glucozei la zaharificare (Yoo și colab., 2018), exploatând această diversitate genetică prin reproducere convențională, ajutată de studii de Asociere la nivel de genom (GWAS) (Porth și colab., 2013; Fahrenkrog și colab., 2017; Liu și colab., 2018), reproducere cu alele rare defecte (BRDA) (Vanholme și colab., 2013a) sau selecție genomică (Yin și colab., 2010; Muchero și colab., 2015; Pawar și colab., 2018; Xie și colab., 2018), este o strategie valoroasă pentru obținerea liniilor care au îmbunătățit eficiența prelucrării lemnului. Odată ce copacii de elită sunt obținuți prin aceste metode de reproducere, ingineria genetică și editarea genetică bazată pe CRISPR a genelor specifice este o cale foarte promițătoare de a îmbunătăți în continuare aceste genotipuri de elită fără a le rupe constituția genetică și fără a trece prin cicluri lungi de reproducere. Având în vedere iminentul accident climatic, nu mai avem timp de pierdut în adoptarea acestor noi tehnici de reproducere în cursa noastră către economia bazată pe bio.
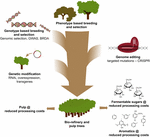
Figura 3. Îmbunătățirea genetică a copacilor forestieri printr-o combinație de instrumente de reproducere. Pentru a accelera îmbunătățirea genetică a copacilor forestieri pentru aplicațiile de celuloză și biorafinare, instrumentele de reproducere clasice și noi trebuie combinate inteligent. Reproducerea clasică implică selecția fenotipică a copacilor pentru cruci controlate, urmată de selecția fenotipică. Odată cu apariția informațiilor despre secvența genomului multor copaci forestieri, au fost dezvoltate noi strategii, cum ar fi selecția genomică, studiile de Asociere la nivel de genom (GWAS) și reproducerea cu alele rare defecte (BRDA) pentru a accelera captarea și îmbogățirea polimorfismelor ADN asociate cu trăsături benefice. Editarea genomului bazată pe CRISPR permite modificarea genomului într-un mod care imită polimorfismele naturale. Modificarea genetică implică integrarea stabilă a ADN-ului străin în arbore pentru a supraproduce (o) enzimă(e) sau a regla în jos (a) genă(e). Combinarea tehnicilor de reproducere clasice și noi este necesară pentru a oferi suficient lemn de înaltă calitate pentru societate.
contribuțiile autorului
toți autorii enumerați au adus o contribuție substanțială, directă și intelectuală la lucrare și au aprobat-o pentru publicare.
finanțare
recunoaștem finanțarea parțială din proiectul IWT-SBO BIOLEUM (Grant Nr.130039) și de către SBO-FISH prin proiectul ARBOREF. AC a primit finanțare din partea FWO și a programului de cercetare și inovare Horizon 2020 al Uniunii Europene în cadrul Acordului de Grant Marie sk, nr.665501. LdV a fost finanțat de Institutul pentru promovarea inovării prin știință și Tehnologie din Flandra (IWT-Vlaanderen) pentru o bursă predoctorală.
Declarație privind conflictul de interese
autorii declară că cercetarea a fost realizată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
mulțumiri
îi mulțumim lui Annick Bleys pentru pregătirea acestui manuscris.
Boerjan, W., Ralph, J. și Baucher, M. (2003). Biosinteza ligninei. Anu. Rev. Plante Biol. 54, 519–546.
PubMed Abstract/Google Scholar
Lapierre, C., Pollet, B., Petit-Conil, M., Toval, G., Romero, J., Pilat, G. și colab. (1999). Modificările structurale ale ligninelor în plopii transgenici cu activitate deprimată de alcool cinamilic dehidrogenază sau acid cafeic o-metiltransferază au un impact opus asupra eficienței pulpării industriale kraft. Physiol De Plante. 119, 153–163.
PubMed Abstract/Google Scholar
Ralph, J. (2006). „Ce face un bun înlocuitor monolignol?, „în știința și tradiția peretelui celular al plantei – biosinteza, structura și funcția, ed. T. Hayashi (Boca Raton, FL: BrownWalker Press).
Google Scholar