eOrganic forfatter:
Dr. Mark schonbeck, Virginia Association for biological farming
abstract
grise er det almindelige navn for flere nært beslægtede sommerårige, der er blevet store ukrudt af grøntsager og rækkeafgrøder i hele USA og store dele af verden. De fleste grise er høje, oprejst til buskede planter med enkle, ovale til diamantformede, alternative blade og tætte blomsterstande (blomsterklynger) bestående af mange små, grønlige blomster. De dukker op, vokser, blomstrer, sætter frø og dør inden for den frostfrie vækstsæson.
grise trives i varmt vejr, tolererer tørke, reagerer på høje niveauer af tilgængelige næringsstoffer og er tilpasset til at undgå skygge gennem hurtig stængelforlængelse. De konkurrerer aggressivt mod varme sæsonafgrøder og reproduceres ved produktiv frøproduktion.
i økologiske produktionssystemer kan grise styres gennem en kombination af:
- rettidig dyrkning, flammelukning og manuel fjernelse
- forældet såbed
- Mulching
- afgrøderotationer, der varierer timing af jordbearbejdning og andre operationer
- dækafgrøder og konkurrencedygtige kontante afgrøder
- foranstaltninger til at forhindre eller minimere produktion af levedygtige frø
introduktion
stort set alle landmænd i Nordamerika kender og kæmper med grise, et udtryk, der dækker flere arter i slægten Amaranthus, herunder:
- redroot grise (A. (A. albus)
- glat Pileurt (A. hybridus)
- Pileurt (A. potelii)
- Palmer amaranth (A. palmeri)
- spiny amaranth (A. spinosus)
- tumble Pileurt (A. albus)
- næsegrus Pileurt (A. blitoides)
- vandhemp (A. spinosus)
- tumble Pileurt (A. albus)
- næsegrus (A. blitoides)
- A. tuberculatus = A. Rudis)
disse varme-elskende sommer enårige dukke efter foråret frost Dato, vokse hurtigt, konkurrere kraftigt mod varme-sæson afgrøder, reproducere ved frø, og dø med efteråret frost. Grise er store ukrudt af varme sæsongrøntsager (2006) og rækkeafgrøder (Sellers et al., 2003).
også kaldet amaranths, grise er hjemmehørende i dele af Nord-og Mellemamerika. Afgrødedyrkning og menneskelig handel har åbnet nye nicher, der gør det muligt for grise at invadere landbrugsøkosystemer i hele Amerika og dele af Europa, Asien, Afrika og Australien. De fleste amaranter fremstiller næringsrige grønne grøntsager eller kornafgrøder, og bevidst plantning til mad har hjulpet nogle ukrudtsarter spredt rundt om i verden. Imidlertid dyrkes ingen af de svineukrudt, der diskuteres her, kommercielt til korn, og moderne amaranth-sorter betragtes ikke som store landbrugsukrudt.
Svineproblemerne er steget i No-till produktionssystemer med konventionelle herbicider, som efterlader ukrudtsfrø på overfladen og vælger til herbicidresistente populationer (Sellers et al., 2003). Imidlertid kan høje svinepopulationer forekomme på økologiske og ikke-økologiske gårde og i konventionelle, bevarings-og No-till-systemer.
beskrivelse og identifikation
grise er lette at genkende, men korrekt identifikation af grisearter kan være vanskelig. To eller flere grisearter forekommer ofte sammen i det samme felt (Fig. 1), kan der forekomme signifikant variation inden for en art, og interspecifikke hybrider forekommer lejlighedsvis (Sellers et al., 2003). Nogle forskere betragter højt vandhemp og almindeligt vandhemp som en enkelt art: A. tuberculatus (Pratt and Clark, 2001). Kansas State University udvidelse har udgivet en fremragende griseidentifikationsvejledning med fotoillustrationer og en nøgle til at skelne modne planter af ni forskellige ukrudt amaranter (Horak et al., 1994).

Figur 1. To grisearter, foreløbigt identificeret som Palmer amaranth (venstre) og glat griseurt (højre), vokser ved kanten af en plastisk mulket seng i økologisk vegetabilsk produktion i Clemson, South Carolina. Foto kredit: Mark Schonbeck, Virginia Association for biologisk landbrug.
nyopståede piggeblomster åbner et par lange, smalle cotyledoner, cirka 0,5 tommer lange med 0,1 tommer brede, efterfulgt af de første ægte blade, som er bredere i omridset (Fig. 2). Planter danner moderat dybe, forgrenede taproots og kan vise en tydelig rødlig farve på rødder, nedre stængler og undersider af blade.

figur 2. I denne flush af sommeren årlige ukrudtsplanter, grise (Amaranthus sp.) kan skelnes ved dets par lange, smalle cotyledoner (frøblade) og på ældre frøplanter ægte blade, der er meget mere bredt ovale i omrids. Foto kredit: Mark Schonbeck, Virginia Association for biologisk landbrug.
de fleste grise vokser til store, oprejst til buskede planter, 2-7 fod i højden, med enkle, blomstrende (stilkede) blade arrangeret skiftevis (enkeltvis) på stilke (Fig. 3a). Bladbladene er generelt ovale til diamantformede og 2-6 inches lange. Næsegrus danner en lav, spredende måtte med mindre (ca.en tomme) blade, der er tydeligt hakket ved spidsen (Fig. 3b).

figur 3. disse glatte grise i tidlig overskrift er omkring fire meter høje. b. næsegrus danner en lav, spreder måtten. Foto kreditter: Mark Schonbeck, Virginia Association for biologisk landbrug.
individuelle griseblomster er små, iøjnefaldende og normalt grønlige i farve. Mandlige og kvindelige blomster bæres på samme plante (de fleste arter) eller separate planter (vandhemp, Palmer amaranth). Hver plante bærer tusinder af blomster i små klynger i bladaksler eller større, ofte forgrenede, tætpakkede pigge ved spidserne af hovedstængler og store grene (Fig. 4). Kvindelige blomster danner enkelt, små, runde, normalt skinnende, mørke rødbrune til sorte frø, omtrent 0,04 tommer i diameter (Fig. 5). Omkring 50.000 – 90.000 frø vejer en ounce.

figur 4. Spiny amaranth (venstre) og glat grise (højre) i blomst. Foto kredit: Mark Schonbeck, Virginia Association for biologisk landbrug.
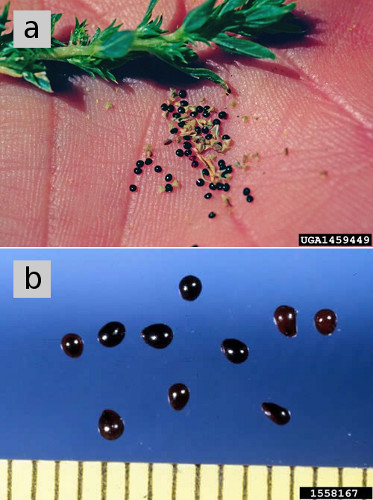
figur 5. a) frø af tumbler. (B) frø af rødrødt gris, forstørret, Viser mørk, skinnende frø frakke af modne frø. Figur kreditter: (a) Steve Duey, Utah State University, Bugwood.org. (b) Ken Chamberlain, Ohio State University, Bugwood.org.
Se tabel 1 nedenfor for en hurtig guide til otte almindelige nordamerikanske grisearter med links til yderligere oplysninger om hver.
| fælles og Videnskabeligt navn | vækstvaner | blomsterstand* | geografisk rækkevidde** | andre Plantekarakteristika | |
|---|---|---|---|---|---|
| redroot gris Amaranthus retrofleksus | oprejst, forgrenet, 2-7 ft | stive, forgrenede terminalspidser, individuelle grene normalt <2 i lang, tykkere end blyant | i hele Nordamerika inklusive Alaska | øvre stilk og blade normalt dækket med fine hår; bladblade Store (6 tommer) på kraftige planter | |
| glat gris Amaranthus hybridus | oprejst, forgrenet, 2-7 ft | bløde, stærkt forgrenede terminalspidser, individuelle grene tyndere end blyant | i hele Nordamerika | svarende til redroot, men meget variabel, mange lokale varianter, kan hybridisere med nært beslægtede arter | |
| palmer amaranth Amaranthus Palmeri | oprejst, forgrenet, 2-10 ft | lang (til 18 tommer), enkle eller sparsomt forgrenede terminalspidser; mandlig blød, kvindelig børstet | sydlige halvdel af USA. Ekstremt hurtig, aggressiv vækst i varme klimaer, mandlige og kvindelige blomster på separate planter; planter glat og hårløs | ||
| Potell Amaranth Amaranthus potellii | oprejst, forgrenet, 2-6 ft | stive, forgrenede terminalspidser, grene 4-8 i lang, tykkere end blyant, holdt tæt på hovedaksen | i hele Nord Amerika |
første ægte blade smallere og mere koniske mod spidsen end redroot eller glat; planten kan være glat eller Behåret | |
| Spiny Amaranth Amaranthus spinosus | oprejst til busket 1-4 ft | slanke, forgrenede terminalspidser for det meste mandlige blomster; aksillære klynger for det meste kvindelige | i hele Nordamerika, men for det meste sydøstlige USA | par stive, skarpe Kurt-in rygsøjler ved bunden af hvert blad; stængler glat, hårløs, ofte rød | |
| vandhemp Amaranthus Rudis eller A. Tuberculatus*** | oprejst, høj 3-10 ft | slanke, enkle eller forgrenede terminalspidser | i hele USA. og sydlige Canada undtagen tørreste områder | mandlige og kvindelige blomster på separate planter; stængler og blade glatte og hårløse; blade ofte længere og smallere end andre arter | |
| næsegrus Pileurt Amaranthus blitoides | næsegrus mat til 3 ft tværs | små, tætte klynger i bladøkser | i hele USA og det sydlige Canada | ||
| Tumble Grisegræs Amaranthus albus | kugleformet busk, 1-3 ft diameter | små, tætte klynger i bladaksler | i hele Nordamerika | modne planter bryder af på jordoverfladen og bæres af vind, spreder frø; stængler hvid til lysegrøn, blade lysegrøn | |
| * små klynger af blomster er normalt til stede i bladaksler af alle Amaranter ** inden for Nordamerika (Canada, USA, Rusland); mange arter er blevet naturaliseret på andre kontinenter. * * * nogle forfattere genkender to arter, almindelig vandhemp (A. rudis) og høj vandhemp (A. tuberculatus); andre betragter dem som underarter eller synonyme. |
|||||
livscyklus, reproduktion, frøspredning, Frødvalen og spiring
grise er frost-ømme sommerårige, der dukker op, vokser, blomstrer og danner modent frø inden for den frostfrie periode. Frøplanter dukker op over en længere periode med store skylninger i slutningen af foråret eller forsommeren (Fig. 6). I de fleste arter finder blomstring og frøudvikling hovedsageligt sted efter sommersolhverv som reaktion på forkortelse af daglængder.

figur 6. En flush af glatte grisefrøplanter på en grøntsagsbedrift i Tidevandsregionen i Virginia, fotograferet den 20.juni 2010, cirka to uger efter fremkomsten. Foto kredit: Mark Schonbeck, Virginia Association for biologisk landbrug.
grise reproducerer helt af frø. En enkelt stor plante kan modne 100.000-600.000 frø og populationer på 0.1-1 planter pr. kvadratfod kan kaste 10.000–45.000 frø pr.kvadratfod eller 0,4-2 milliarder pr. acre (Massinga et al., 2001; sælgere et al., 2003). Denne produktive frøproduktion gør grise særligt vanskelige at håndtere, da vellykket modning af kun en plante pr.10.000 nye frøplanter kan gøre det muligt for grisebestanden at stige flere gange fra et år til det næste.
grise begynder typisk at blomstre og kaste pollen (antese) omkring seks uger efter fremkomsten (vej), selvom blomster kan forekomme så tidligt som 3 vej eller så sent som 9 vej (Huang et al., 2000; Keeley, et al., 1987; Shrestha og Svanton, 2007). 1-2 uger efter, at blomsterknopper først bliver synlige for det blotte øje.rapporterede tidsintervaller fra bestøvning til dannelse af levedygtige frø spænder fra 7-12 dage i vandhemp (Bell and Tranel , 2010) til 6 uger i feltpopulationer af rødrødt grise i Ontario (Shrestha og Svanton, 2007). I Californien dannede Palmer amaranth levedygtige frø 2-6 uger efter blomstring (Keeley et al., 1987). Frø bliver levedygtige på omtrent samme tid, som de udvikler deres modne mørkebrune eller sorte farve.
reproduktiv udvikling accelereres ved at forkorte daglængden efter sommersolhverv i feltpopulationer (Keeley et al., 1987), og fortsætter hurtigere på kort (~12 timer) end i længere (cirka 14 timer) fotoperioder i et vækstkammer (Huang et al., 2000). Selvom de fleste frøproduktion forekommer i sensommeren og det tidlige efterår, er der fundet nogle modne frø i glatte grisefrøhoveder ved sommersolhverv i Virginia (personlig observation).
griseplanternes evne til at blive rykket op med rode eller afskåret ved blomstring for at fuldføre frømodning er ikke undersøgt. Men i upstate NY York, 2-4-tommer fragmenter af Potell amaranth blomsterstande liggende på jordoverfladen viste sig at indeholde sorte frø 3 uger efter ukrudtet blev disked ned ved blomstring (Charles Mohler, Cornell University, pers. commun.). Tilsyneladende, hvis bestøvning finder sted, før svinekødene trækkes eller hugges, eksisterer der et vist potentiale for levedygtig frøproduktion.
Grisefrø spredes til nye steder ved vanding eller oversvømmelsesvand, gødning og jord, der klæber fast på fodtøj, traktordæk eller jordbearbejdningsværktøjer. Derudover spreder tumble-grise aktivt frø, når modne planter bryder af og bevæger sig med vinden.
Grisefrø har flere dvalemekanismer, så frø produceret i en given sæson spirer på forskellige tidspunkter i løbet af de næste mange år og derved forbedrer ukrudtets langsigtede vedholdenhed (Egley, 1986). Nyskurte grisefrø er for det meste sovende og bliver mindre i det følgende forår. Spiring fremmes af høje temperaturer (95 liter F), diurnalt svingende temperaturer (f. eks. 85-95 liter F dag, ~ 70 liter f nat) og undertiden lys (Guo og Al-Khatib, 2003; Schonbeck og Egley, 1980 og 1981 Steckel et al., 2004).
grise kommer lettest ud fra den øverste 0,5–1,0 tomme af jordprofilen, hvor Få kommer ud af frø placeret dybere end en tomme (Mohler og Di Tommaso, upubliceret). Frøene kræver tilstrækkelig fugt og god frø–jord kontakt for at absorbere fugt og spire. Mere dybt nedgravede frø forbliver sovende og levedygtige i flere år og spirer, når de bringes til overfladen ved Jordbearbejdning eller dyrkning. Selvom flushes af fremkomst ofte følger forberedelse eller dyrkning af såbed, stigende svineproblem i agronomiske afgrøder er tilskrevet udbredt vedtagelse af no-till og minimum-Jordbearbejdning, der efterlader ukrudtsfrø for nylig ved eller nær jordoverfladen (Sellers et al., 2003).
vækstvaner og indvirkning på afgrøder
grise har C4 fotosyntetisk vej, som giver en evne til at vokse hurtigt ved høje temperaturer og høje lysniveauer, at tolerere tørke og at konkurrere aggressivt med varmsæsongrøntsager om lys, fugt og næringsstoffer. Vækst er relateret til kumulative Vækstgradsdage med en basetemperatur på 50 liter F (Shrestha og Svanton, 2007; Horak og Loughin, 2000); således vokser svinekøds meget hurtigere i varme klimaer end i nordlige regioner med køligere somre.
oprejste svinearter kan hurtigt vælte korte afgrøder som broccoli eller snapbønne. I højere afgrøder som majs reagerer grise på baldakinskygge ved at øge stammevæksten og indsætte blade højere på planten og derved opfange en større brøkdel af tilgængeligt lys (Massinga et al., 2003; McLachlan et al, 1993). 10 fod række, der opstår med majs eller sojabønne, kan forårsage betydelige udbyttetab (Klingman og Oliver, 1994; Knesevic et al., 1994; Massinga et al., 2001) grise, der opstår flere uger efter, at afgrøden er opstået, har meget mindre effekt på udbyttet.
grise reagerer meget på næringsstoffer, især nitratformen af nitrogen (N) (Sortsja og Brandt, 2008; Teyker et al., 1991). Befrugtning forbedrer både ukrudtsbiomasse og frøproduktion. Derudover kan nitrat stimulere spiring af grisefrø (Egley, 1986). En barkflis af bælgplanter dække afgrøderester er blevet observeret for at øge gris fremkomsten i nogle år (Fig. 7), sandsynligvis som følge af hurtig mineralisering af legume N (Teasdale og Mohler, 2000).

Figur 7. I dette feltforsøg konkurrerer en flush af grise mod broccoli plantet no-till I dræbt Behåret vetch (forgrund), mens broccoli plantet i dræbt rug eller rug–vetch er relativt fri for grise (baggrund). Hurtig n mineralisering fra afgrøderester med hele bælgplanten stimulerede tilsyneladende spiring og vækst af grise. Foto kredit: Mark Schonbeck, Virginia Association for biologisk landbrug.
fordi de små frø har minimale næringsreserver, er svinekødplanter oprindeligt mere afhængige af let tilgængelige næringsstoffer fra jorden, især fosfor (P) og kalium (K), end større frøede planter såsom majs, bønner og cucurbitae (Hoveland et al., 1976; Mohler, 1996). I undersøgelser udført på organisk (muck) jord i Florida var glat grise og spiny amarant mindre lydhør end salat til P-niveauer, og en bandpåføring af p-gødning forbedrede afgrødens evne til at konkurrere mod disse ukrudt (Santos et al., 1997; Shrefler et al., 1994).
grise er skyggeintolerante, og væksten og reproduktionen af individer, der opstår under en tung afgrøde baldakin, reduceres væsentligt. Hurtig stængelforlængelse gør det imidlertid muligt for grise at undslippe skygge i mange beskæringssituationer. Sensæson grise, der bryder igennem etableret cucurbit, tomat, peber og andre grøntsager, kan fremme afgrødesygdom ved at reducere luftcirkulationen, forstyrre høsten og sætte mange tusinde frø (Fig. 8).

figur 8. Grisen opstod flere uger efter plantning af græskar og påvirkede ikke udbyttet. Ved afslutningen af afgrødehøsten, imidlertid, hver ukrudt modnet tusinder af frø, og vil foretage en tung indbetaling i ukrudtsfrøbanken, medmindre de fjernes straks. Foto Kredit: Mark Schonbeck, Virginia Association for biologisk landbrug.
Svineukrudt rapporteres at være vært for skadedyrsnematoder (Meloidogyne spp.) og mange vegetabilske afgrødepatogener, herunder svampe, der forårsager tidlig rødme i kartoffel og tomat (Alternaria solani), salatdråbe (Sclerotinia sclerotiorum) og sydlig rødme (Sclerotium rolfsii) i en lang række afgrøder. Virale patogener såsom agurkmosaikvirus og tomatplettet visnevirus kan også overføres fra grise (Mohler og DiTommaso, upubliceret).
grise er blevet fokus for biokontrolindsatsen med svampepatogener og plantefodrende insekter, selvom der endnu ikke er nogen biokontrolprodukter til rådighed for landmændene. Amaranth loppebille (Disonycha glabrata) forekommer i store dele af USA (Tisler, 1990), lever af griseblade (Fig. 9), og kan blive en betydelig naturlig fjende af grise i nogle områder, herunder Floyd County, Virginia (personlig observation). Det kontrollerer dog normalt ikke ukrudtet og lever lejlighedsvis af nogle grøntsagsplanter.

figur 9. Den Amarante loppebille lever af grisegræs løv, og det er blevet observeret at forårsage betydelig afblødning og reducere ukrudtskraft i nogle dele af Virginia. Dette insekt kan lejlighedsvis blive et skadedyr i sukkerroer og chard ved at fodre på frøplanter. Foto kredit: Mark Schonbeck, Virginia Association for biologisk landbrug.
Ledelse
økologiske landmænd forvalter grise ved at udnytte deres sårbarhedspunkter. De små frø har minimale næringsreserver; således kan frøplanter kun komme ud af frø placeret inden for en tomme af jordoverfladen og er straks afhængige af jorden for let tilgængelige næringsstoffer. Transplanterede og storfrøede afgrøder har betydelige næringsreserver og kan få en konkurrencefordel i forhold til svinekødplanter, hvis der anvendes næringskilder med langsom frigivelse.
de sarte frøplanter dræbes let ved afskæring, rodning, begravelse eller varme. Rettidig flamme lugning eller dyrkning med en hvilken som helst af en række redskaber kan banke en flush af Pileurt kimplanter. Emerging grise er også modtagelig for skygge og fysisk hindring af mulch. En feltundersøgelse ved Beltsville, Maryland dokumenterer den større følsomhed af grise til undertrykkelse med organiske mulches i forhold til flere andre almindelige ukrudt: redroot grise > lammekvarteret > giant rævhale > velvetleaf (Teasdale og Mohler, 2000).
rettidig handling er afgørende, da grise hurtigt bliver sværere at dræbe, når de vokser højere end en tomme og udvikler fire eller flere ægte blade (Fig. 10). I kølige klimaer kan grisefrøplanter forblive sårbare over for dyrkning i op til 4 vej (væver og Mcvilliams, 1980); men i varmere klimaer kan de vokse til 2-4 tommer inden for 2 vej (Sellers et al., 2003).

Figur 10. Grisefrøplanten til højre er på det sårbare stadium, hvor den let kan dræbes ved lav dyrkning eller flammning eller blokeres af barkflis. Når grise vokser så stort som frøplanten til venstre, bliver det sværere at dræbe, hvilket kræver mere kraftig dyrkning. Foto Kredit: Mark Schonbeck, Virginia Association for biologisk landbrug.
Grisebestande tilpasser sig let til produktionssystemer og kontroltaktik. For eksempel viser frøspredningsresponser adaptive ændringer i forskellige afgrøderotationer (Brainard et al., 2007), og udbredt herbicidresistens er blevet rapporteret i flere arter (Fugate, 2009; Volenberg et al., 2007). Således vil afhængighed af et enkelt styringsværktøj eller den samme strategi år efter år sandsynligvis give faldende afkast over tid.
når de anvendes i kombination, kan de nedenfor beskrevne fremgangsmåder give effektiv håndtering af svin i organiske systemer.
dyrkning og flamme ukrudt
Overvåg afgrøder regelmæssigt for ukrudtsopkomst. Dyrk, når grise er i cotyledon-scenen, eller før de når en tomme i højden, og arbejder så tæt på afgrøderækken som praktisk. Når afgrøden er tilstrækkeligt etableret, sæt kultivatorer til at flytte en tomme eller deromkring jord i rækker for at begrave små ukrudt. Overfladelaget af løs, tør jord, der efterlades ved dyrkning (støvbarkflis), afskrækker yderligere spiring af grise. Undgå at komprimere jorden igen, da komprimering kan fremme en anden flush af fremkomst (Fig. 11).

Figur 11 dyrkning efterlod en støvbarkflis omkring disse unge courgetteplanter og derved afskrækker spiring af grise og andre småfrøede ukrudt. Imidlertid, fodtrafik komprimerede jorden nok til at genoprette frø–jordkontakt nær overfladen, derved tillader ukrudtsfrø at absorbere fugt, spire, og vokse i fodsporene. Foto Kredit: Mark Schonbeck, Virginia Association for biologisk landbrug.
Flammelukning kan fjerne grise og andre bredbladede frøplanter lige før afgrødens fremkomst. Det bruges ofte til langsomt startende afgrøder som gulerod, sukkerroer og pastinak. Fordi flammende normalt ikke dræber græsplanter, anbefales det ikke, hvor græs udgør en betydelig del af ukrudtsfloraen.
klipning og græsning
når afgrøden er for stor til at dyrke med traktor, klipper, klipper eller trækker landmænd ofte ukrudt i gyder for at opretholde luftcirkulationen omkring afgrøden, Letter høsten og forhindrer ukrudtsformering. Dette skal gøres, før svineblomster åbnes (inden for få dage efter, at blomsterhoveder først bliver synlige) for at forhindre levedygtig frødannelse.
nogle landmænd klipper gyder mellem brede rækker eller plastmulkede senge med en push-plæneklipper eller line-trimmer som et jordbesparende alternativ til dyrkning. To rettidige klipninger inden baldakinlukning har givet tilstrækkelig kontrol mellem rækken af kæmpe rævehale, svinekødog ambrosia i sojabønne plantet i en 30-tommers rækkeafstand (Donald, 2000).
de fleste grise er meget velsmagende for husdyr. Modne frø passerer imidlertid uskadt gennem dyrenes fordøjelseskanaler, og gødning er en berygtet kilde til grisefrø. Således bør grise græsses, mens de stadig er vegetative. Bemærk også, at det nationale økologiske Program kræver et 120-dages interval mellem gødningsaflejringer fra græssende dyr og den næste høst af fødevareafgrøder.
Mulching
Mulching kan være en effektiv kontrol taktik for grise i vegetabilsk produktion. En organisk barkflis, såsom 3-4 tommer halm eller hø (~5-10 tons/ac), der påføres inden for en dag efter dyrkning af en etableret afgrøde, kan reducere den efterfølgende svineveds fremkomst med 90%. Alternativt kan en syntetisk barkflis, såsom sort plast, lægges inden afgrødeplantning og gyde ukrudt kontrolleret ved dyrkning, klipning, organisk barkflis eller dækafgrøde. Bemærke: Hvis plast eller anden syntetisk mulch bruges til økologiske afgrøder, skal den fjernes fra marken i slutningen af høsten eller vækstsæsonen.
organisk no-till transplantation af tomat og andre sommergrøntsager til rullekrympede eller klippede vinterafdækningsafgrøder kan kontrollere let til moderat grisebestand. Rug rester frigiver naturlige plantevæksthæmmere (allelokemikalier), der undertrykker grise og nogle andre årlige ukrudt (Barnes og Putnam, 1983; Putnam et al., 1983) uden at påvirke transplanterede grøntsager.
næringsstof-og Fugtighedsstyring
brug langsomt frigivende kilder til N og andre afgrødenæringsstoffer, og undgå udsendelse af hurtigere frigivelsesmaterialer som blodmel og knoglemel, som kan give grise springet på afgrøden. Til tunge foderstoffer som broccoli eller spinat, der har brug for nogle hurtige n -, bånd-eller sidekjole-materialer inden for eller i nærheden af afgrøderækken ved begyndelsen af hurtig afgrødevækst.
brug drypvanding i træk til at give vand og flydende organisk gødning direkte til afgrøden uden fodring og vanding mellem ukrudt. Dryppelinjer under jorden kan give afgrøden fugt og lade jordoverfladen være tør og derved minimere ukrudtsfremkomst inden for rækken.
sædskifte, Planteskemaer og forældet såbed
Planlæg sædskifte og planlæg feltoperationer for at forstyrre svineveds livscyklus. Undgå at give en åben niche (bar jord) år efter år for grisebryst fremkomst i slutningen af foråret til forsommeren. Alternativ varme-og kølige sæson grøntsager. Overvej at forsinke forberedelse af såbed til en sommergrøntsag indtil efter tidspunktet for peak griseved fremkomst. Efter flere års intensiv vegetabilsk produktion skal du dreje marken til flerårig sod (f.eks.
Hvis grisebestanden er høj (Fig. 12), Forbered et uaktuelt såbed i det sene forår for at trække ukrudtsfrøbanken ned. Till eller dyrk, rull eller kultipack jorden for at forbedre frø–jordkontakt og derved fremme ukrudtsspiring. Drys vand, hvis jorden er tør. Gentag dyrkning efter behov. Lige før afgrødeplantning eller afgrødeopkomst skal du bruge lav dyrkning og lade overfladen være løs for at modvirke yderligere ukrudtsspiring. Den endelige flush kan også dræbes af flamme, hvis græs ukrudt er få eller fraværende.

Figur 12. Et tæppe af spiny amaranth kimplanter stammer fra en stor ukrudts frøbank. En forældet såbed eller dyrket Brak er nødvendig for at bringe denne situation under kontrol. Foto kredit: Mark Schonbeck, Virginia Association for biologisk landbrug.
Afgrødekonkurrence og Dækbeskæring
med god ukrudtsbekæmpelse i den tidlige sæson kan kraftige afgrøder som tomat, sød kartoffel og vinterkvas tåle senere voksende grise. Imidlertid, afgrødekonkurrence kontrollerer muligvis ikke ukrudt, på grund af deres skygge-undgåelsesrespons og evne til at bryde igennem afgrødebaldakinen gennem hurtig stængelforlængelse.
konkurrencedygtige sommerdækafgrøder som boghvede, sorghum–sudangrass, koær og foder sojabønne bruges ofte til at undertrykke ukrudt mellem forår og efterår vegetabilske afgrøder. I Florida, koær, sunnhemp, eller velvetbean dækafgrøder podet med høje hastigheder reduceret, men eliminerede ikke glat vækst af grise (Collins et al., 2008).
når du bruger sommerdækafgrøder til at bekæmpe svinekødsfrø, frø ved høje hastigheder (1,5–2 gange normalt) og brug gode såningsmetoder til at opnå et ukrudtsundertrykkende dækafgrødestativ. Kombiner koær, foder sojabønne eller andet sommerbælg med et højt græs som perlehirse eller sorghum–sudangrass for at udvikle en baldakin, der er både høj og tæt. Se afgrøden nøje; hvis der vokser en betydelig mængde grise med det, skal du straks afslutte afgrøden, når ukrudtsblomsthoveder først vises.
håndtering af Grisefrøbanken
fordi svinekødsfrø producerer frø så produktivt, er det afgørende at minimere den årlige frøregn på jorden. Selvom streng ukrudtsbekæmpelse i seks år kan reducere grisefrøbanken med 99%, giver afslappende ukrudtsbekæmpelse frøantal mulighed for at komme sig til næsten oprindelige niveauer inden for tre år (1984). Grise, der opstår efter en afgrødes minimale ukrudtsfri periode, reducerer muligvis ikke afgrødens udbytte, men det skal trækkes eller skæres inden blomstring for at forhindre dannelse af modne frø.
det kan betale sig at gå marker med modne afgrøder for at trække eller hugge store ukrudt ud; små, forkrøblede grise under en afgrødebaldakin danner kun et lille antal frø. Hvis der allerede er dannet blomsterhoveder, skal du fjerne afskårne eller rodede griseplanter fra marken. Hvis griseplanter allerede har dannet frø, skal du bemærke, at mange af frøene forbliver i hovedet indtil vinteren. Derfor kan fjernelse af ukrudt i det tidlige efterår stadig reducere grisefrøregnen markant.
i tilfælde af at der opstår en kraftig grisefrøregn, anbefaler nogle ukrudtsforskere inversion Jordbearbejdning for at flytte frø til en dybde, hvorfra de ikke kan komme ud (Mohler og Di Tommaso, upubliceret). Selvom 5-14% af redroot grise og vandhemp frø har overlevet 9-12 års begravelse på 8 tommer dybde i Nebraska (Burnside et al., 1996), har andre rapporteret, at grisefrø er ret kortvarige (3-4 år) i jorden i mere fugtige regioner som Mississippi og Illinois (Buhler og Hartsler, 2001; Egley og Vilhelm, 1990; Steckel et al., 2007). Moldboard pløjning er blevet rapporteret at øge grise fremkomsten, hvis ukrudtspopulationer er lave, men for at mindske fremkomsten, hvis populationer er høje som følge af en nylig frø regn (1984).
Når du bruger inversion Jordbearbejdning til at styre en tung frøaflejring, pløjer moldboard marken en gang og undgår derefter dyb jordbearbejdning i de næste mange år for at lade nedgravede frø miste levedygtighed.
denne artikel er en del af en serie, der diskuterer den invasive familie af grise. For mere information, se følgende artikler:
- Ukrudtsprofil: grise (Amaranthus spp.)
- Redroot Pigweed (Amaranthus retroflexus)
- Powell Amarant (Amaranthus powellii)
- Tornede Amarant (Amaranthus spinosus)
- Palmer Amarant (Amaranthus palmeri)
- Glat Pigweed (Amaranthus hybridus)
- Tumble Pigweed (Amaranthus albus)
- Næsegrus pigweed (Amaranthus blitoides)
- Common Waterhemp (Amaranthus rudis) og Høj Waterhemp (A. tuberculatus)
Referencer Citeret
- Barnes, J. P., A. R. Putnam. 1983. Rugrester bidrager til ukrudtsundertrykkelse i afgrødesystemer uden Jordbearbejdning. Tidsskrift for kemisk økologi 9: 1045-1057. (Tilgængelig online på: http://dx.doi.org/10.1007) (bekræftet 10 September 2012).
- Bell, M. S. og P. J. Tranel. Tidskrav fra bestøvning til frø modenhed i vandhemp (Amaranthus tuberculatus). Ukrudtsvidenskab 58: 167-173. (Tilgængelig online på: http://dx.doi.org/10.1614/WS-D-09-00049.1) (bekræftet 10 September 2012).R. E. Og R. N. Brandt. 2008. Kvælstofgødning Sats virkninger på ukrudt konkurrenceevne er arter afhængige. Ukrudtsvidenskab 56: 743-747. (Tilgængelig online på: http://dx.doi.org/10.1614/WS-08-065.1) (bekræftet 10 September 2012).
- Brainard, D. C., A. DiTommaso og C. A. Mohler. 2007. Intraspecifik variation i frøegenskaber ved Potell amaranth (Amaranthus potellii) fra levesteder med kontrasterende afgrøderotationshistorier. Ukrudtsvidenskab 55: 218-226. (Tilgængelig online på: http://dx.doi.org/10.1614/WS-06-134.1) (bekræftet 10 September 2012).
- Buhler, D. D. og R. G. Hartler. 2001. Fremkomst og vedholdenhed af frø af fløjlblad, almindelig vandhemp, uldig kopgræs og kæmpe rævehale. Ukrudtsvidenskab 49: 230-235. (Tilgængelig online på: http://dx.doi.org/10.1614/0043-1745(2001)049%5B0230:EAPOSO%5D2.0.CO; 2) (bekræftet 10.September 2012).han er en af de mest kendte og mest kendte mennesker i verden. 1996. Frø levetid af 41 ukrudtsarter begravet 17 år i det østlige og vestlige Nebraska. Ukrudtsvidenskab 44: 74-86. (Tilgængelig online på: http://www.jstor.org/stable/4045786) (bekræftet 10 September 2012).Collins, A. S., C. A. Chase og C. M. Hutchinson. 2008. Optimale tætheder af tre bælgplanter til undertrykkelse af glat grise (Amaranthus hybridus). Ukrudtsvidenskab 56: 753-761. (Tilgængelig online på: http://dx.doi.org/10.1614/WS-07-101.1) (bekræftet 10.September 2012).
- Donald, 2000. Mellem-rækken græsslåning + i-rækken band-anvendt herbicid til ukrudtsbekæmpelse i glycin maks. Ukrudtsvidenskab 48: 487-500. (Tilgængelig online på: http://www.jstor.org/stable/4046280) (bekræftet 10 September 2012).
- Egley, G. H. 1986. Stimulering af ukrudtsfrøspiring i jord. Anmeldelser af Ukrudtsvidenskab 2: 67-89.
- Egley, G. H. og R. D. 1990. Tilbagegang af ukrudtsfrø og frøplanteopkomst over fem år som påvirket af jordforstyrrelser. Ukrudtsvidenskab 38: 504-510. (Tilgængelig online på: http://www.jstor.org/stable/4045064) (bekræftet 10 September 2012).
- Fugate, L. 2009. Grise får landmænd til at genoverveje landbrugsmetoder. University of Arkansas Division of Agriculture Cooperative udvidelse Service Nyheder-oktober 2009.Guo, P. og K. Al-Khatib. 2003. Temperaturpåvirkninger på spiring og vækst af rødrodsved (Amaranthus retrofleksus), Palmer amaranth (A. palmeri) og almindelig vandhemp (A. rudis). Ukrudtsvidenskab 51: 869-875. (Tilgængelig online på: http://dx.doi.org/10.1614/P2002-127) (bekræftet 10 September 2012).Horak, M. J. og T. M. Loughin. 2000. Vækstanalyse af fire Amaranthusarter. Ukrudtsvidenskab 48: 347-355. (Tilgængelig online på: http://dx.doi.org/10.1614/0043-1745(2000)048%5B0347:GAOFAS%5D2.0.CO; 2) (bekræftet 10.September 2012).
- Horak, M. J., D. E. Peterson, D. J. Chessman og L. M. voks. 1994. Griseidentifikation: en Pictoral guide til de almindelige grise på de store sletter. 12 s. (Tilgængelig online på: http://www.ksre.ksu.edu/bookstore/pubs/S80.pdf) (bekræftet 6 August 2013).Hoveland, C. S., G. A. Buchanan og M. C. Harris. 1976. Reaktion af ukrudt på jord fosfor og kalium. Ukrudtsvidenskab 24: 194-201. (Tilgængelig online på: http://www.jstor.org/stable/4042586) (bekræftet 10.September 2012).Huang, A. Shrestha, M. Tollenar, M. Deen, H. Rahimian og C. J. Svanton. 2000. Effekt af fotoperiode på den fænologiske udvikling af rødrodsved (Amaranthus retrofleksus L.). Canadisk Tidsskrift for plantevidenskab 80: 929-938.Keeley, P. E., C. H. Carter og R. J. Thullen. 1987. Indflydelse af plantedato på vækst af Palmer amaranth (Amaranthus palmeri). Ukrudtsvidenskab 35: 199-204. (Tilgængelig online på: http://www.jstor.org/stable/4044391) (bekræftet 10 September 2012).
- Klingman, T. E. Og L. R. Oliver. 1994. Palmer amaranth (Amaranthus palmeri) interferens i sojabønner (glycin maks). Ukrudtsvidenskab 42: 523-527. (Tilgængelig online på: http://www.jstor.org/stable/4045448) (bekræftet 10.September 2012).det er en af de mest populære ting i verden. 1994. Der er ingen grund til at tro, at det er nødvendigt at tage hensyn til, om det er nødvendigt. Ukrudtsvidenskab 42: 568-573. (Tilgængelig online på: http://www.jstor.org/stable/4045456) (bekræftet 10.September 2012).
- Massinga, R. A., R. S. Currie, M. J. Horak og J. Boyer, Jr.2001. Indblanding af Palmer amaranth i majs. Ukrudtsvidenskab 49: 202-208. (Tilgængelig online på: http://dx.doi.org/10.1614/0043-1745(2001)049%5B0202:IOPAIC%5D2.0.CO; 2) (bekræftet 10.September 2012).
- Massinga, R. A., R. S. Currie og T. P. Trooien. 2003. Vandforbrug og let aflytning under Palmer amaranth (Amaranthus palmeri) og majs konkurrence. Ukrudtsvidenskab 51: 523-531. (Tilgængelig online på: http://dx.doi.org/10.1614/0043-1745(2003)051%5B0523:WUALIU%5D2.0.CO; 2) (bekræftet 10.September 2012).han er en af de mest kendte i verden, og han er en af de mest kendte i verden. 1993. Effekt af majs-induceret skygge på tørstof ophobning, distribution og arkitektur af rødrod Pileurt (Amaranthus retrofleksus). Ukrudtsvidenskab 41: 568-573. (Tilgængelig online på: http://www.jstor.org/stable/4045424) (bekræftet 10 September 2012).
- Mohler, C. A. 1996. Økologiske baser til kulturel kontrol af årlige ukrudt. Tidsskrift for produktion landbrug 9: 468-474..
- Mohler, C. A. og A. DiTommaso. Ikke offentliggjort. Administrer ukrudt på din gård: en Guide til økologiske strategier. Institut for afgrøde-og Jordvidenskab, Cornell University. Udkast til præpublikation, version 5.1. Offentliggørelse forventes i 2012.Pratt, D. B. Og L. G. Clark. 2001. Amaranthus rudis og A. tuberculatus-en art eller to? Tidsskrift for Torrey Botanical Society 128: 282-296. (Tilgængelig online på: http://www.jstor.org/stable/3088718) (bekræftet 10 September 2012).
- Putnam, A. R., J. DeFrank og J. P. Barnes. 1983. Udnyttelse af allelopati til ukrudtsbekæmpelse i årlige og flerårige dyrkningssystemer. Tidsskrift for kemisk økologi 9: 1001-1010. (Tilgængelig online på: http://dx.doi.org/10.1007) (bekræftet 10 September 2012).det er en af de mest populære og mest populære destinationer i verden. 1997. Effekt af fosforfrugtbarhed på konkurrencemæssige interaktioner mellem glat grisegræs (Amaranthus hubridus), spiny amaranth (Amaranthus spinosus) og almindelig purslane (Portulaca oleracea) med salat. Ukrudt videnskab Society of America Abstracts 37: 54.
- Schonbeck, M. V. og G. H. Egley. 1980 Redroot grise (Amaranthus retrofleksus) frøspiring svar på eftermodning, temperatur, ethylen og nogle andre miljømæssige faktorer. Ukrudtsvidenskab 28: 543-548. (Tilgængelig online på: http://www.jstor.org/stable/4043277) (bekræftet 10 September 2012).
- Schonbeck, M. V. og G. H. Egley. 1981. Ændringer i følsomheden af Amaranthus retrofleksus L. frø til ethylen under præinkubation. II. virkninger af skiftende temperatur og nedgravning i jord. Plante, celle og Miljø 4: 237-242. (Tilgængelig online på: http://dx.doi.org/10.1111/1365-3040.ep11611005) (bekræftet 10 September 2012).E. E. og R. L. Simdahl. 1984. Ukrudtsfrø falder i kunstvandet jord efter seks års kontinuerlig majs og herbicider. Ukrudtsvidenskab 32: 76-83. (Tilgængelig online på: http://www.jstor.org/stable/4043886) (bekræftet 10 September 2012).
- sælgere, B. A., R. J. Smeda, Johnson, J. A. Kendig og M. R. Ellersieck. 2003. Sammenlignende vækst af seks Amaranthus arter i Missouri. Ukrudtsvidenskab 51: 329-333. (Tilgængelig online på: http://dx.doi.org/10.1614/0043-1745(2003)051%5B0329:CGOSAS%5D2.0.CO; 2) (bekræftet 10.September 2012).det er en af de mest populære og mest populære måder at gøre det på. 1994. Fosforfrugtbarhedens virkning på konkurrencen mellem salat (Lactuca sativa) og spiny amaranth (Amaranthus spinosus). Ukrudtsvidenskab 42: 556-560. (Tilgængelig online på: http://www.jstor.org/stable/4045454) (bekræftet 10 September 2012).
- Shrestha, A. og C. J. Svanton. 2007. Parameterisering af den fænologiske udvikling af udvalgte årlige ukrudt under ikke-beskårne markforhold. Ukrudtsvidenskab 55: 446-454. (Tilgængelig online på: http://dx.doi.org/10.1614/WS-06-176.1) (bekræftet 10.September 2012).Steckel, L. E., C. L. Sprague, E. V. Stoller og L. M. voks. 2004. Temperatureffekter på spiring af ni Amaranthusarter. Ukrudtsvidenskab 52: 217-221. (Tilgængelig online på: http://dx.doi.org/10.1614/WS-03-012R) (bekræftet 10 September 2012).Steckel, L. E., C. L. Sprague, E. V. Stoller, L. M. voks og F. V. Simmons. 2007. Jordbearbejdning, beskæringssystem og jorddybdeeffekter på almindelig vandhemp (Amaranthus rudis) frøbankens vedholdenhed. Ukrudtsvidenskab 55: 235-239. (Tilgængelig online på: http://dx.doi.org/10.1614/WS-06-198) (bekræftet 10 September 2012).
- Teasdale, J. R. og C. L. Mohler. 2000. Det kvantitative forhold mellem ukrudtsfremkomst og mulches fysiske egenskaber. Ukrudtsvidenskab 48: 385-392. (Tilgængelig online på: http://dx.doi.org/10.1614/0043-1745(2000)048%5B0385:TQRBWE%5D2.0.CO; 2) (bekræftet 10.September 2012).
- Teyker, R. H., H. D. og R. A. Liebl. 1991. Majs og svinebjørnsrespons på kvælstofforsyning og-form. Plante og jord 135: 287-292. (Tilgængelig online på: http://dx.doi.org/10.1007) (bekræftet 10.September 2012).
- Tisler, A. M. 1990. Fodring i grisebjælken, Disonycha glabrata Fab. (Coleoptera: Chrysomelidae), på Amaranthus retrofleksus. Virginia Journal of Science 41: 243-245.Volenberg, D. S., A. G. Hager og P. J. Tranel. 2007. Svar på nutidig og historisk vandhemp (Amaranthus tuberculatus) tiltrædelser af glyphosat. Ukrudtsvidenskab 55: 327-333. (Tilgængelig online på: http://dx.doi.org/10.1614/WS-06-121) (bekræftet 10 September 2012).
- væver, S. E. og E. L. Mcvilliams. 1980. Biologi af canadiske ukrudt. 44. Amaranthus retrofleksus L, A. A. hybridus L. Canadian Journal of Plant Science 60: 1215-1234.T. M. 2006. Ukrudtsundersøgelse-sydlige stater. Underafdeling af grøntsager, frugter og nødder. Forløbet af det sydlige Ukrudtsvidenskabssamfund 59: 260-277. (Tilgængelig online på: http://www.swss.ws/wp-content/uploads/docs/Southern%20Weed%20Survey%202006%20Vegetables%20and%20Fruits.pdf) (bekræftet 10.September 2012).