1-1. Los glóbulos blancos del sistema inmunitario derivan de precursores en la médula ósea
Todos los elementos celulares de la sangre, incluidos los glóbulos rojos que transportan el oxígeno, las plaquetas que desencadenan la coagulación de la sangre en los tejidos dañados y las células blancas de la sangre del sistema inmunitario, derivan en última instancia de las mismas células precursoras o precursoras: las células madre hematopoyéticas de la médula ósea. Como estas células madre pueden dar lugar a todos los tipos diferentes de células sanguíneas, a menudo se las conoce como células madre pluripotentematopoyéticas. Inicialmente, dan lugar a células madre de potencial máslimitado,que son los progenitores inmediatos de los glóbulos rojos, las plaquetas y las dos categorías principales de glóbulos blancos. Los diferentes tipos de células sanguíneas y sus relaciones de linaje se resumen en la Fig. 1.3. Nos ocuparemos aquí de todas las células derivadas del progenitor linfoide común y del progenitor mieloide, aparte de los megacariocitos y los glóbulos rojos.

Figura 1.3
Todos los elementos celulares de la sangre, incluidos los linfocitos del sistema inmunitario adaptativo, surgen de células madre hematopoyéticas en la médula ósea. Estas células pluripotentes se dividen para producir dos tipos más especializados de células madre, un linfoide común(más…)
El progenitor mieloide es el precursor de los granulocitos,macrófagos, células dendríticas y mastocitos del sistema inmunitario. Los macrófagos son uno de los tres tipos de fagocitos del sistema inmunitario y se distribuyen ampliamente en los tejidos corporales, donde desempeñan un papel fundamental en la inmunidad innata.Son la forma madura de monocitos, que circulan en la sangre y se diferencian continuamente en macrófagos al migrar a los tejidos. Las células dendríticas se especializan en recuperar antígeno y mostrarlo para su reconocimiento por los linfocitos. Las células endríticas inmaduras migran de la sangre para residir en los tejidos y son tanto fagocíticas como macropinocíticas, ingiriendo grandes cantidades del líquido extracelular circundante. Al encontrar un patógeno, maduran rápidamente y migran a los ganglios linfáticos.
Los mastocitos, cuyos tumores no están bien definidos, también se diferencian en los tejidos. Se encuentran principalmente cerca de pequeños vasos sanguíneos y, cuando se activan, liberan sustancias que afectan a la permeabilidad vascular. Aunque son más conocidos por su papel en la reducción de las respuestas alérgicas, se cree que desempeñan un papel en la protección de las superficies de la mucosa contra los patógenos.
Los granulocitos se llaman así porque tienen gránulos densamente teñidos en su citoplasma; también a veces se les llama polimorfonucleococitos debido a sus núcleos de forma extraña. Hay tres tipos de granulocitos, todos los cuales son de vida relativamente corta y se producen en mayor número durante las respuestas inmunitarias, cuando salen de la sangre para migrar a sitios de infección o inflamación. Los neutrófilos, que son la tercera célula fagocítica del sistema inmunitario, son el componente celular más numeroso e importante de la respuesta inmune innata: las deficiencias hereditarias en la función de los neutrófilos conducen a una infección bacteriana abrumadora, que es mortal si no se trata. Se cree que los eosinófilos son importantes principalmente en la defensa contra las infecciones parasitarias, porque su número aumenta durante una infección parasitaria. La función de los basófilos es probablemente similar y complementaria a la de los eosinófilos y los mastocitos; discutiremos las funciones de estas células en el capítulo 9 y su papel en la inflamación alérgica en el Capítulo 12. Las células del linaje mieloide se muestran en la Fig. 1.4.
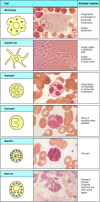
Figura 1.4
Células mieloides en inmunidad innata y adaptativa. Las células del linaje mieloide realizan varias funciones importantes enla respuesta inmunitaria. Las celdas se muestran esquemáticamente en la columna izquierda en la forma en que se representarán a lo largo de la columna (más…)
El progenitor linfoide común da lugar a los linfocitos, de los que se ocupará la mayor parte de este libro. Hay dos tipos principales de linfocitos: linfocitos B o células B, que cuando se activan se diferencian en células plasmáticas que secretan anticuerpos; y linfocitos T o células T,de los cuales hay dos clases principales. Una clase se diferencia en la activación de las células T intocitotóxicas, que matan las células infectadas con virus, mientras que la segunda clase de células T se diferencia en células que activan otras células, como las células B y los macrófagos.
La mayoría de los linfocitos son células pequeñas, sin rasgos distintivos, con pocos orgánulos citoplásmicos y gran parte de la cromatina nuclear inactiva, como se muestra en su estado condensado (Fig. 1.5). Esta apariencia es típica de las células inactivas y no es sorprendente que, tan recientemente como a principios de la década de 1960,los libros de texto pudieran describir estas células, ahora el foco central de la inmunología, sin función conocida. De hecho, estos pequeños linfocitos no tienen actividad funcional hasta que encuentran antígeno, que es necesario para desencadenar su proliferación y la diferenciación de sus características funcionales especializadas.

Figura 1.5
Los linfocitos son en su mayoría células pequeñas e inactivas. El panel izquierdo muestra una micrografía ligera de un linfocito pequeño rodeado de glóbulos rojos. Nótese la cromatina condensada del núcleo, que indica poca actividad transcripcional, la ausencia relativa(más…)
Los linfocitos son notables por ser capaces de montar una respuesta inmunitaria específica contra prácticamente cualquier antígeno extraño. Esto es posible porque cada linfocito individual madura con una variante única de un prototipo de receptor de antígenos, de modo que la población de linfocitos T y B tiene colectivamente un gran repertorio de receptores que son muy diversos en sus sitios de unión a antígenos. El receptor de antígeno de células B (BCR) es una forma unida a la membrana del anticuerpo que las células B secretarán después de la activación y diferenciación a las células plasmáticas. Las antibodimoléculas como clase se conocen como inmunoglobulinas, generalmente acortadas a Ig, y el receptor de antígenos de los linfocitos B es, por lo tanto, también conocido como inmunoglobulina de membrana (mIg).El receptor de antígenos de células T (TCR) está relacionado con la inmunoglobulina, pero es bastante distinto de ella, ya que está especialmente adaptado para detectar antígenos derivados de proteínas extrañas o patógenos que han entrado en las células huésped. Describiremos en detalle las estructuras de estos receptores de antígenos linfocitos en los capítulos 3, 4 y 5, y la forma en que se crea su diversidad de sitios de unión a medida que se desarrollan los linfocitos en el capítulo 7.
Un tercer linaje de células linfoides, llamadas células asesinas naturales, carecen de receptores antígenespecíficos y forman parte del sistema inmunitario innato. Estas células circulan en la sangre como grandes linfocitos con gránulos citotóxicos distintivos (Fig. 1.6). Son capaces de reconocer y matar algunas células anormales,por ejemplo, algunas células tumorales y células infectadas por virus, y se cree que son importantes en la defensa inmune innata contra los patógenos intracelulares.

Figura 1.6
Células asesinas naturales (NK). Se trata de grandes células granulares similares a linfocitos con funciones importantes en la inmunidad innata. Aunque carecen de receptores específicos de antígenos, pueden detectar y atacar ciertas células infectadas por virus.Fotografía cortesía de N. Rooney (más información…)