Introducción
La interacción exitosa con nuestro entorno sensorial requiere un intrincado equilibrio de dos mecanismos de selección de atención: el de arriba hacia abajo y el de abajo hacia arriba. Dirigirse al pasillo de productos de su supermercado local con el objetivo de recoger pocos ingredientes necesarios para la ensalada de mango, implica el despliegue de un sistema de atención voluntario dirigido a objetivos o de arriba hacia abajo, de modo que busque activamente todos los ingredientes necesarios entre la multitud de opciones de productos. Sin embargo, si escucha el timbre de un teléfono celular, lo más probable es que capture su atención e interrumpa su búsqueda. Tal interrupción ocurre de manera ascendente, o impulsada por estímulos, por lo que una mera prominencia del estímulo, el hecho de que el anillo es diferente de otros sonidos en su entorno, lo considera digno de selección. El escenario descrito subraya la importancia de la selección de comportamiento dirigida a objetivos e impulsada por estímulos, y apunta a un delicado equilibrio que debe existir entre los dos sistemas de atención para evitar la «visión de túnel» por un lado y la incapacidad total para enfocarse en el otro.
Selección de arriba hacia abajo y de abajo hacia arriba: Comportamiento
Varias décadas de investigación conductual se han dedicado a demostrar que la distribución de la atención puede ser controlada por las intenciones del observador, así como por la prominencia del estímulo físico. Gran parte de la evidencia conductual para la asignación de atención de arriba hacia abajo y de abajo hacia arriba se ha revisado ampliamente en otros lugares (Johnston y Dark, 1986; Egeth y Yantis, 1997). En resumen, los estudios que demuestran los efectos del control de atención de arriba hacia abajo muestran que la atención se puede asignar con éxito a ubicaciones espaciales, características, objetos, etc., tras la presencia de señales exógenas o endógenas (Eriksen y Hoffman, 1972; Posner, 1980; Posner et al., 1980), o expectativas establecidas por el conocimiento previo o por contingencias del estímulo (Shaw, 1978; Moore y Egeth, 1998; Geng y Behrmann, 2002, 2005; Shomstein y Yantis, 2004a; Drummond y Shomstein, 2010). La evidencia que apoya la asignación de atención de abajo hacia arriba se ha basado en varios paradigmas de captura de atención, en los que los participantes se involucran en una búsqueda de arriba hacia abajo y su atención se desvía a los estímulos irrelevantes para la tarea, lo que demuestra que la atención es capturada por elementos individuales de características (unique item; Yantis y Jonides, 1990; Theeuwes, 1991; Folk et al., 2002) y abruptos (Yantis y Jonides, 1984; Theeuwes, 1991; Koshino et al., 1992; Juola et al., 1995).
Mientras que la mayoría de los estudios iniciales se concentraron en demostrar evidencia para la selección de atención de arriba hacia abajo y de abajo hacia arriba, los estudios más recientes cambiaron su enfoque a examinar cómo interactúan los dos sistemas de selección de atención. Esta línea de investigación está alimentada por observaciones que, para seleccionar de manera efectiva la información relevante para la tarea (por ejemplo, los ingredientes para la ensalada), uno debe inhibir activamente la información irrelevante para la tarea que de otro modo desviaría la atención de la tarea en cuestión. La otra cara de esta lógica, es que cuanto menos se enfoque en la información relacionada con la tarea, más captura se producirá. Se ha demostrado experimentalmente que el estado de atención del observador predice qué tipo de información, y en qué medida, finalmente captará la atención(Folk et al., 1992, 2002; Bacon y Egeth, 1994; Gibson y Kelsey, 1998). Por ejemplo, Folk et al. (2002) mostraron que al buscar una letra roja, un observador será capturado más fácilmente por un estímulo irrelevante en la periferia si ese estímulo es rojo, o coincide con la plantilla objetivo de alguna manera. Dado que los ajustes de control de arriba hacia abajo del observador están configurados para buscar una característica roja, es probable que cualquier estímulo que sea rojo capte la atención y potencialmente interfiera con el control de arriba hacia abajo. Por lo tanto, con una tarea de captura, las estrategias de búsqueda atencional se pueden distinguir entre sí variando los niveles de similitud entre las propiedades de estímulo del objetivo y los distractores. Cuanto más similar sea el objetivo al distractor, más difícil será para el observador evitar la captura.
El papel del Lóbulo Parietal en la Selección de Arriba hacia Abajo y de Abajo hacia Arriba: Varias técnicas de neuroimagen proporcionaron pruebas sólidas de la implicación de la corteza parietal en la orientación de arriba hacia abajo y de abajo hacia arriba, y las pruebas se revisaron ampliamente en otros lugares (Corbetta y Shulman, 2002, 2011; Behrmann et al., 2004). Se ha demostrado que las áreas más comúnmente activadas siguiendo señales de arriba hacia abajo para atender a ubicaciones, características u objetos particulares se encuentran a lo largo de las partes dorsales de la corteza parietal. Tales áreas incluyen lóbulo parietal inferior( IPL), regiones dorsomediales denominadas lóbulo parietal superior (SPL), así como regiones más medias a lo largo del giro precuneo (Yantis et al., 2002; Giesbrecht et al., 2003; Liu et al., 2003; Yantis y Serences, 2003; Figura 1). Se han demostrado varias tareas de arriba hacia abajo para involucrar con éxito regiones dorsales de la corteza parietal, a saber, aquellas que involucran el espacio (Kastner et al., 1999; Corbetta et al., 2000; Hopfinger et al., 2000; Shomstein y Behrmann, 2006; Chiu y Yantis, 2009; Greenberg et al., 2010), así como los cambios de atención no espaciales (Giesbrecht et al., 2003; Yantis y Serences, 2003; Shomstein y Yantis, 2004b, 2006; Tamber-Rosenau et al., 2011).
FIGURA 1.
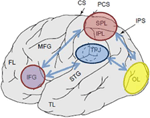
la Figura 1. Representación esquemática de puntos de referencia anatómicos relevantes proyectados en la superficie lateral del cerebro humano. El lóbulo parietal superior (LPS) y el lóbulo parietal inferior (LPI) son regiones dentro de la parte dorsal de la corteza parietal que sirven de orientación atencional de arriba hacia abajo. La unión temporo-parietal (TPJ, por sus siglas en inglés) es una región dentro de la corteza parietal ventral que sirve de orientación atencional de abajo hacia arriba. Se cree que ambos, SPL y TPJ, provocan señales de control responsables de las modulaciones atencionales posteriores observadas en regiones sensoriales, en este caso modulando (etiquetadas con flechas azules oscuras) la actividad evocada visualmente en el lóbulo occipital (OL). Además, se cree que las áreas a lo largo del giro frontal inferior (IFG) y la unión frontal inferior (IFJ) sirven como áreas de convergencia para el control atencional impulsado por estímulos y de arriba hacia abajo (marcadas con flechas bidireccionales de color azul claro).
la Figura 1. Representación esquemática de puntos de referencia anatómicos relevantes proyectados en la superficie lateral del cerebro humano. El lóbulo parietal superior (LPS) y el lóbulo parietal inferior (LPI) son regiones dentro de la parte dorsal de la corteza parietal que sirven de orientación atencional de arriba hacia abajo. La unión temporo-parietal (TPJ, por sus siglas en inglés) es una región dentro de la corteza parietal ventral que sirve de orientación atencional de abajo hacia arriba. Se cree que ambos, SPL y TPJ, provocan señales de control responsables de las modulaciones atencionales posteriores observadas en regiones sensoriales, en este caso modulando (etiquetadas con flechas azules oscuras) la actividad evocada visualmente en el lóbulo occipital (OL). Además, se cree que las áreas a lo largo del giro frontal inferior (IFG) y la unión frontal inferior (IFJ) sirven como áreas de convergencia para el control atencional impulsado por estímulos y de arriba hacia abajo (marcadas con flechas bidireccionales de color azul claro).
En una tarea típica destinada a involucrar la asignación de atención de arriba hacia abajo, a los individuos se les muestran dos flujos de presentación visual en serie rápida (RSVP) ubicados periféricamente y se les indica inicialmente que monitoreen un flujo para obtener una señal (por ejemplo, un dígito entre el flujo de letras). La identidad de la señal indica si el sujeto debe mantener la atención en la corriente actual o cambiar la atención a la otra corriente(Yantis et al., 2002; Yantis y Serences, 2003). En estos paradigmas se observan dos hallazgos importantes. La primera tiene que ver con una mayor activación dentro de las regiones sensoriales que representan la ubicación atendida en el momento (por ejemplo, una mayor actividad dentro de las regiones visuales primarias izquierdas cuando se asiste a la corriente de confirmación de asistencia derecha). Este hallazgo proporciona evidencia firme de que los participantes están atendiendo a un lugar específico y que la atención modula la fuerza de la respuesta sensorial (ver Figura 1; Moran y Desimone, 1985; O’Craven et al., 1997). El segundo hallazgo tiene que ver con la observación de que las regiones dorsales del lóbulo parietal se activan selectivamente por cambios de atención de arriba hacia abajo. Se observa que el curso temporal de actividad de SPL/IPL es de naturaleza transitoria, lo que sugiere que esta área de la corteza parietal es la fuente de una breve señal de control de atención para cambiar los estados de atención de una manera descendente(Yantis et al., 2002).
Varios estudios de IRMF han documentado que la captura de atención de abajo hacia arriba, mediada por la prominencia y/o relevancia del estímulo, está subservida por la unión temporoparietal (TPJ; Figura 1). Por ejemplo, cuando los sujetos atienden y monitorean un cambio en un estímulo visual o auditivo, presentado simultáneamente, se mejora la activación de las regiones TPJ del lóbulo parietal. Además de la aparente sensibilidad a estímulos relevantes, el TPJ también se activa en respuesta a eventos potencialmente novedosos (inesperados o infrecuentes) cuando un organismo se involucra en un contexto de comportamiento neutral o cuando se involucra en una tarea (Marois et al., 2000; Downar et al., 2002; Serences et al., 2005; Corbetta et al., 2008; Asplund et al., 2010; Diquattro y Geng, 2011; Geng y Mangun, 2011). Esta activación ocurre independientemente de la modalidad (auditiva, táctil y visual) en la que se entrega la entrada, lo que refleja la naturaleza multisensorial del TPJ (pero ver Downar et al., 2001).
En una tarea típica que examina el mecanismo neuronal de la captura de atención de abajo hacia arriba, a los participantes se les presenta un flujo de RSVP de elementos en el centro de la pantalla y se les pide que identifiquen un objetivo predefinido (por ejemplo, identificar la letra roja presentada dentro de un flujo de RSVP de blancos no objetivos). Una parte de los ensayos contiene un distractor saliente irrelevante para la tarea presentado en varios intervalos de tiempo antes del inicio del objetivo, mientras que otros ensayos contienen solo el distractor saliente (es decir, sin el objetivo). Los ensayos de «distractor objetivo» se utilizan para evaluar el alcance de la captura, mostrando que el distractor irrelevante para la tarea es de hecho sobresaliente, lo que produce una disminución en la precisión del objetivo. Los ensayos de «distractor aislado» se utilizan para análisis adicionales, ya que dichos ensayos permiten examinar la actividad inducida al distractor saliente sin contaminación de los procesos relacionados con el objetivo. De esos paradigmas surgen varias conclusiones importantes. En primer lugar, cuando los distractores se separan espacialmente de la ubicación objetivo, los distractores de captura se acompañan de una mayor actividad cortical en las regiones correspondientes de la corteza sensorial (por ejemplo, corteza visual organizada retinotópicamente; consulte la Figura 1). Tales resultados proporcionan una fuerte evidencia de que durante la captura, la atención espacial se captura de hecho en la ubicación espacial ocupada por el distractor (Serences et al., 2005). En segundo lugar, las regiones ventrales de la corteza parietal, principalmente dentro del TPJ, se activan selectivamente por cambios de atención involuntarios de abajo hacia arriba. Al igual que la actividad dentro de la SPL para la orientación de arriba hacia abajo, el curso de tiempo de la actividad observada sobre TPJ es de naturaleza transitoria, lo que sugiere que esta región es la fuente de una breve señal de control de atención para cambiar la atención de una manera ascendente.
Cabe señalar que, si bien esta revisión se centra en abordar las funciones cognitivas de la corteza parietal posterior, otras regiones, especialmente las que se encuentran dentro de la corteza frontal, también se reclutan para la asignación de atención de arriba hacia abajo y de abajo hacia arriba. Tales regiones incluyen la corteza frontal ventral( VFC), los campos oculares frontales (FEF), la unión frontal inferior (FIP) y el giro frontal inferior (IFG; Corbetta y Shulman, 2002, 2011; Serences et al., 2005; Asplund et al., 2010; Diquattro y Geng, 2011).
El papel del lóbulo Parietal en la Selección de Arriba hacia Abajo y de Abajo hacia Arriba: Neuropsicología
Históricamente, los investigadores se basaron críticamente en estudios neuropsicológicos de pacientes con negligencia hemispacial (un trastorno de la asignación espacial de atención al hemiespacio izquierdo) para obtener información sobre las funciones cognitivas asociadas con el lóbulo parietal. En la literatura neuropsicológica clásica, la corteza parietal, en su totalidad, se consideró generalmente el sitio de la lesión primaria para negligencia hemispacial. Esta visión, elaborada en detalle por los primeros investigadores (Critchley, 1953; McFire y Zangwill, 1960; Piercy, 1964) reconoció claramente la asociación entre la lesión parietal y el descuido resultante. Esta perspectiva se mantuvo en gran medida a lo largo de la década de 1980, cuando Posner y sus colegas (1984) utilizaron el paradigma de la señal visuoespacial encubierta para mostrar que el daño al lóbulo parietal produce un déficit en la operación de «desconexión» (retrayendo la atención de un lugar y desplazándola a otro) cuando el objetivo es contralateral a la lesión. Sin embargo, a pesar de este gran avance en la comprensión de la base neuronal de la atención y específicamente el papel de «desconexión» de la corteza parietal, sus hallazgos asumen un único sitio cortical (corteza parietal) y una única capacidad funcional («desconexión»). En contraste con este enfoque más monolítico del cerebro (corteza parietal) y el comportamiento (desconexión atencional), el trabajo reciente de comportamiento y neuroimagen (revisado anteriormente y en otros lugares) sugiere que tanto la región cortical como el comportamiento atencional asociado pueden subdividirse en perfiles cualitativamente diferentes.
Dada la segregación de las redes corticales en procesos de arriba hacia abajo y de abajo hacia arriba, una predicción obvia es que el daño a las porciones superiores del lóbulo parietal (subsumiendo SPL) debería producir un déficit en la orientación atencional dirigida a objetivos, mientras que el daño a las porciones inferiores del lóbulo parietal (subsumiendo TPJ) resultaría en un déficit asociado con la captura de atención impulsada por estímulos. En la medida en que estas correspondencias de comportamiento cerebral han sido exploradas en la literatura neuropsicológica, esta predicción no se mantiene obviamente. Por ejemplo, los síntomas clínicos de negligencia hemispacial están fuertemente asociados con daño a las porciones inferiores del lóbulo parietal, que incluye TPJ, en lugar de a porciones superiores como el LPS (Friedrich et al., 1998; Shomstein et al., 2010; Corbetta y Shulman, 2011). Esto está un poco en desacuerdo con la literatura de neuroimagen, que sugiere que el papel de la TPJ está en la captación de la atención, más que en la orientación voluntaria de la atención, el dominio en el que los pacientes descuidados parecen tener la mayor dificultad. Para complicar aún más las cosas, se ha observado que las lesiones que involucran exclusivamente LPS, solo raramente producen evidencia clínica de negligencia (Vallar y Perani, 1986). Otro estudio reciente con pacientes con lesiones centradas principalmente en TPJ y STG pero con LPS preservado, Corbetta et al. (2005) mostraron que la negligencia espacial, así como su recuperación, se asoció con la restauración de la actividad en las regiones ventral temporoparietal y dorsal parietal (ver Corbetta y Shulman, 2011 para una revisión). Aunque sus conclusiones son interesantes y emocionantes, este último estudio no diferencia la contribución relativa de las vías dorsal y ventral a diferentes tipos de atención, ya que los pacientes solo se probaron en una variante de la tarea de atención espacial encubierta Posner, tarea que se cree que involucra la orientación de atención de arriba hacia abajo y de abajo hacia arriba.
Para distinguir entre el control atencional orientado a objetivos y la captura atencional sobresaliente y para examinar su mapeo en el SPL y el TPJ, respectivamente, un estudio reciente adoptó dos paradigmas de comportamiento, cada uno dirigido a una de estas formas de atención (Shomstein et al., 2010). Para examinar la integridad de la orientación de atención de arriba hacia abajo en los pacientes, se utilizó una tarea de arriba hacia abajo que requería que los participantes cambiaran la atención espacial entre las corrientes de RSVP separadas espacialmente (una tarea que se ha utilizado con éxito para demostrar la activación del LPS en estudios de IRMF (Yantis et al., 2002)). De manera similar, para examinar las capacidades de orientación de atención de abajo hacia arriba de los pacientes, una variante de Folk et al. (2002) se empleó el paradigma de captura contingente en el que los participantes detectaron objetivos que aparecían en la fijación mientras que los singletes de color irrelevantes para la tarea se destellaban en la periferia. La medida en que los distractores irrelevantes de la tarea interfieren con la tarea de detección central se utilizó entonces como una medida de captura de atención de abajo hacia arriba (Bacon y Egeth, 1994; Folk et al., 2002).
Las predicciones fueron las siguientes: los pacientes con lesiones en las porciones superiores del lóbulo parietal (que afectan a la LPS) deben verse afectados en la tarea de orientación de atención de arriba hacia abajo (con un rendimiento conservado en la tarea de captura), mientras que los pacientes con lesiones en las porciones inferiores del lóbulo parietal (que afectan a la TPJ) deben verse afectados en la tarea de captura (con un rendimiento preservado en la tarea de arriba hacia abajo). Una doble disociación de esta forma no solo da fe de los componentes independientes de la atención, sino que también sugiere que dichos componentes de atención están mediados por mecanismos neuronales independientes. Ocho pacientes con negligencia visuoespacial fueron reclutados para el estudio y completaron dos tareas, aprovechando la orientación atencional impulsada por estímulos o dirigida a objetivos. En función de su perfil conductual, los pacientes se clasificaron en grupos y se exploró la superposición de lesiones (Figura 2A). Los pacientes que presentaron dificultades con la orientación atencional dirigida a objetivos, cuantificada por el índice de atención de arriba hacia abajo (Figura 2B), presentaron superposición de lesiones centradas en porciones superiores del lóbulo parietal (subsumiendo LPS) con lóbulo parietal inferior preservado (TPJ). Los pacientes con lesiones superpuestas centradas en las porciones inferiores del lóbulo parietal (que subsumen TPJ), pero que no sufrieron LPS, se desempeñaron normalmente en la tarea de orientación dirigida al objetivo, al tiempo que permanecieron inmunes a la captura de atención (Figura 2C). Los hallazgos de este estudio sugieren claramente que la LPS y la TPJ son regiones anatómicas que se reclutan necesariamente para fines de orientación de arriba hacia abajo y de abajo hacia arriba y que el daño a la LPS y la TPJ conduce a trastornos de orientación de arriba hacia abajo y de abajo hacia arriba, respectivamente.
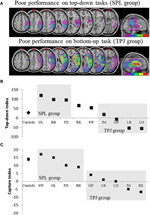
la Figura 2. Resultados del estudio neuropsicológico destinado a investigar la contribución relativa de SPL y TPJ a la orientación de arriba hacia abajo y de abajo hacia arriba. (A) Solapamientos de lesiones (solapamiento mínimo púrpura; solapamiento máximo rojo) para pacientes agrupados por déficits de comportamiento en la orientación de atención de arriba hacia abajo, etiquetados como grupo de LPS (panel superior); y pacientes agrupados por déficits de comportamiento en la orientación de abajo hacia arriba, etiquetados como grupo de TPJ (panel inferior). (B) Desempeño conductual en la tarea de arriba hacia abajo, resumido con un «Índice de arriba hacia abajo» que cuantifica las diferencias entre los desplazamientos espaciales de arriba hacia abajo realizados de izquierda a derecha y viceversa. Los controles y el grupo lesionado por TPJ muestran eficiencias similares en la ejecución de cambios espaciales, mientras que los pacientes con lesiones por LPS muestran una eficiencia disminuida. Los datos de control de grupo y de pacientes individuales (etiquetados con las iniciales del paciente) se representan en la abscisa. C) El» índice de captura » es una medida de la atención de abajo hacia arriba y cuantifica la medida en que los distractores irrelevantes de la tarea captan la atención lejos de la tarea. Los controles y el grupo lesionado por SPL muestran valores de captura similares, de modo que ambos grupos son capturados por los distractores irrelevantes de la tarea. El grupo lesionado por TPJ muestra un índice de captura muy reducido (fallo de captura). Nótese que los pacientes fueron colocados en el grupo de LPS o TPJ en función del comportamiento, en lugar de en función de la lesión, por lo tanto, nótese la consistencia con la que los pacientes terminan en el grupo correspondiente.
Interacción Entre la Selección de arriba hacia Abajo y de Abajo hacia Arriba
Aunque aparentemente existe una fuerte asociación entre la orientación dirigida por objetivos y el LPS y la orientación impulsada por estímulos y el TPJ, datos de Shomstein et al. (2010) un estudio de pacientes sugiere que estos dos sistemas no son totalmente independientes. Esta conclusión está respaldada por el hallazgo de que los pacientes con daño por LPS exhibieron un patrón de desempeño etiquetado como «hiper captura.»A diferencia de los controles, para los que solo el distractor de color del objetivo captó la atención (lo que llevó a una menor precisión del objetivo), los distractores de color irrelevantes también demostraron distraer a los pacientes con lesión por LPS. Además, mientras que para los controles la atención fue capturada por distractores solo cuando precedieron al inicio del objetivo, para los pacientes con lesiones de LPS la atención fue capturada incluso por distractores presentados simultáneamente con el objetivo. Este patrón de desempeño se puede explicar mediante el siguiente marco: El SPL es responsable de la orientación de arriba hacia abajo de la atención que incluye determinar los aspectos de los estímulos que son relevantes para la tarea (por ejemplo, search for red target; Corbetta y Shulman, 2002; Serences et al., 2005). Este conjunto de atención restringe el TPJ, de modo que el mecanismo de captura de atención mediado por el TPJ solo se activa por la información relevante de la tarea (por ejemplo, distractores rojos que captan la atención y distractores grises que no captan la atención cuando se busca un objetivo rojo). La ausencia de SPL impide el establecimiento de un conjunto de atención relevante para la tarea y, por lo tanto, cualquier estímulo, relevante para la tarea o no, se considera importante, por lo que capta la atención (por ejemplo, distractor irrelevante para la tarea que capta la atención para el grupo SPL) indiscriminadamente.
Se ha sugerido que SPL y TPJ podrían interactuar de al menos una de dos maneras posibles. La primera posibilidad es que el TPJ sirva como un sistema de alerta que detecta estímulos relevantes para el comportamiento, pero carece de la alta resolución espacial, por lo que cuando se detecta un estímulo relevante para el comportamiento, su ubicación precisa es suministrada por el SPL que almacena mapas espaciales (Kastner et al., 1999; Wojciulik y Kanwisher, 1999; Bisley y Goldberg, 2003; Silver et al., 2005). Una posibilidad hipotética relacionada es que el mecanismo de captura (que incluye TPJ) actúa como un interruptor de la actividad cognitiva en curso cuando se presenta un estímulo conductual relevante (Corbetta y Shulman, 2002, 2011). El patrón de actividad de «hiper-captura» observado en pacientes con LPS preservado pero lesionado proporciona evidencia adicional para la hipótesis de que el TPJ emite una señal de control que termina la tarea en cuestión, sirviendo así como un disyuntor (Corbetta y Shulman, 2002; Serences et al., 2005). Otros estudios recientes de neuroimagen que emplean varios paradigmas han proporcionado evidencia adicional de una relación interactiva entre la orientación de atención de arriba hacia abajo y la de abajo hacia arriba, y posteriormente para la relación entre SPL y TPJ (Serences et al., 2005; Asplund et al., 2010; Diquattro y Geng, 2011).
Mientras que la evidencia de una interacción entre los dos sistemas de atención y los dos sustratos de atención (SPL y TPJ) es fuerte, lo que no está claro es si esta interacción es directa entre SPL y TPJ o si se logra a través de otras regiones intermedias. Como se mencionó anteriormente, las redes de orientación de atención de arriba hacia abajo y de abajo hacia arriba involucran varias regiones dentro de la corteza frontal, por lo que es razonable suponer que la convergencia entre los dos sistemas podría lograrse a través del lóbulo frontal. Dos estudios recientes que investigan la interacción entre la selección de atención de arriba hacia abajo y de abajo hacia arriba proporcionaron evidencia para la FIP y la IFG como posibles sitios de convergencia entre la selección impulsada por estímulos y la dirigida a objetivos (Asplund et al., 2010; Diquattro y Geng, 2011). La FIP y la IFG parecen ser candidatos ideales para dicha interacción dada su participación general en la atención y el control cognitivo, así como su participación en la selección espacial y no espacial (Koechlin et al., 2003; Brass et al., 2005).
El papel del Lóbulo Parietal en la Selección de Arriba hacia Abajo y de Abajo hacia Arriba: Fisiología
Si bien el énfasis de esta revisión se ha puesto predominantemente en estudios humanos, se ha obtenido una gran riqueza de conocimientos sobre la participación de la corteza parietal en la orientación atencional a partir de investigaciones de fisiología de monos (ver revisión reciente de Bisley y Goldberg, 2010). Sin embargo, cuando se trata de examinar las contribuciones relativas de diferentes regiones dentro de la corteza parietal a la orientación atencional de arriba hacia abajo y de abajo hacia arriba, la literatura sobre fisiología de monos se queda corta. La razón principal de esto es que dentro de la corteza de los monos no parece haber evidencia de la misma segregación de control de arriba hacia abajo y de abajo hacia arriba. En cambio, el área intraparietal lateral (LABIO) que originalmente se pensó que estaba involucrada en la planificación de sacadas (Gnadt y Andersen, 1988) está involucrada en la atención visual y actúa como un mapa de prioridades en el que los estímulos externos se representan de acuerdo con su prioridad de comportamiento derivada de manera descendente o ascendente (Colby y Goldberg, 1999; Bisley y Goldberg, 2003, 2010; Balan y Gottlieb, 2006; Ipata et al., 2006; Buschman y Miller, 2007; Gottlieb y Balan, 2010).
Conclusión
Aunque se sabe mucho menos sobre la corteza parietal humana que sobre la corteza homóloga del mono, estudios recientes, empleando métodos de neuroimagen y neuropsicológicos, han comenzado a dilucidar distinciones funcionales y estructurales cada vez más finas. Esta revisión se centró en estudios neuroimágenes y neuropsicológicos recientes que dilucidan los roles cognitivos de las regiones dorsal y ventral de la corteza parietal en la orientación de atención de arriba hacia abajo y de abajo hacia arriba, y en la interacción entre los dos mecanismos de asignación de atención.
Declaración de conflicto de Intereses
El autor declara que la investigación se realizó en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Reconocimientos
Este trabajo fue apoyado por la subvención EY021644 de los Institutos Nacionales de Salud.Asplund, C. L., Todd, J. J., Snyder, A. P., and Marois, R. (2010). Un papel central para la corteza prefrontal lateral en la atención dirigida a objetivos y impulsada por estímulos. NAT. Neurociencia. 13, 507–512.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Bacon, W. F., and Egeth, H. E. (1994). Anulando la captura atencional impulsada por estímulos. Percepto. Psicófisis. 55, 485–496.
Pubmed Abstract | Pubmed Texto Completo
Balan, P. F., and Gottlieb, J. (2006). Integración de entrada exógena en un mapa dinámico de prominencia revelado por la atención perturbadora. J. Neurocirujano. 26, 9239–9249.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Behrmann, M., Geng, J. J., and Shomstein, S. (2004). Corteza parietal y atención. Curr. Opin. Neurobiol. 14, 212–217.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Bisley, J. W., and Goldberg, M. E. (2003). Actividad neuronal en el área intraparietal lateral y atención espacial. Science 299, 81-86.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Bisley, J. W., and Goldberg, M. E. (2010). Atención, intención y prioridad en el lóbulo parietal. Annu. Reverendo Neurocirujano. 33, 1–21.
Resumen de Pubmed / Texto Completo de Pubmed/Texto completo cruzado
Brass, M., Derrfuss, J., Forstmann, B., and von Cramon, D. Y. (2005). El papel de la zona de unión frontal inferior en el control cognitivo. Trends Cogn. Sci. 9, 314–316.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Buschman, T. J., and Miller, E. K. (2007). Control de atención de arriba hacia abajo versus de abajo hacia arriba en las cortezas prefrontal y parietal posterior. Science 315, 1860-1862.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Chiu, Y. C., y Yantis, S. (2009). Una fuente de control cognitivo independiente del dominio para conjuntos de tareas: cambiar la atención espacial y cambiar las reglas de categorización. J. Neurocirujano. 29, 3930–3938.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Colby, C. L., y Goldberg, M. E. (1999). Espacio y atención en la corteza parietal. Annu. Reverendo Neurocirujano. 22, 319–349.
Pubmed Abstract | Pubmed Texto Completo | CrossRef Texto Completo
Corbetta, M., Kincade, J. M., Ollinger, J. M., McAvoy, M. P., y Shulman, G. L. (2000). La orientación voluntaria está disociada de la detección del objetivo en la corteza parietal posterior humana. NAT. Neurociencia. 3, 292–297.
Pubmed Abstract | Pubmed Texto Completo | CrossRef Texto Completo
Corbetta, M., Kincade, M. J., Lewis, C., Snyder, A. Z., y Sapir, A. (2005). Base neuronal y recuperación de déficits de atención espacial en negligencia espacial. NAT. Neurociencia. 8, 1603–1610.
Pubmed Abstract | Pubmed Texto Completo | CrossRef Texto Completo
Corbetta, M., Patel, G., y Shulman, G. L. (2008). The reorienting system of the human brain: from environment to theory of mind (en inglés). Neurona 58, 306-324.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Corbetta, M., y Shulman, G. L. (2002). Control de la atención dirigida a objetivos e impulsada por estímulos en el cerebro. NAT. Reverendo Neurocirujano. 3, 201–215.
Pubmed Abstract | Pubmed Texto Completo | CrossRef Texto Completo
Corbetta, M., y Shulman, G. L. (2011). Redes de atención y abandono espacial. Annu. Reverendo Neurocirujano. 34, 569–599.
Pubmed Abstract / Pubmed Full Text/CrossRef Full Text
Critchley, M. (1953). Los Lóbulos Parietales. Londres, Reino Unido: Hafner Press.
Resumen de Pubmed / Texto completo de Pubmed
Diquattro, N. E., and Geng, J. J. (2011). El conocimiento contextual configura las redes de control de atención. J. Neurocirujano. 31, 18026–18035.
Pubmed Abstract | Pubmed Texto Completo | CrossRef Texto Completo
Downar, J., Crawley, A. P., Mikulis, D. J., y Davis, K. D. (2001). El efecto de la relevancia de la tarea en la respuesta cortical a cambios en estímulos visuales y auditivos: un estudio de IRMF relacionado con eventos. Neuroimagen 14, 1256-1267.
Pubmed Abstract | Pubmed Texto Completo | CrossRef Texto Completo
Downar, J., Crawley, A. P., Mikulis, D. J., y Davis, K. D. (2002). Una red cortical sensible a la prominencia de estímulos en un contexto conductual neutral a través de múltiples modalidades sensoriales. J. Neurofisiol. 87, 615–620.
Resumen de Pubmed / Texto completo de Pubmed
Drummond, L., y Shomstein, S. (2010). Atención basada en objetos: cambio o incertidumbre? Atten. Percepto. Psicófisis. 72, 1743–1755.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Egeth, H. E., y Yantis, S. (1997). Atención visual: control, representación y curso del tiempo. Annu. Reverendo Psychol. 48, 269–297.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Eriksen, C. W., and Hoffman, J. E. (1972). Características temporales y espaciales de la codificación selectiva a partir de pantallas visuales. Percepto. Psicófisis. 12, 201–204.
Folk, C. L., Leber, A. B., and Egeth, H. E. (2002). ¡Te hizo parpadear! La captura atencional contingente produce un parpadeo espacial. Percepto. Psicófisis. 64, 741–753.
Pubmed Abstract / Pubmed Full Text
Folk, C. L., Remington, R. W., and Johnston, J.C. (1992). La orientación encubierta involuntaria depende de los ajustes de control de atención. J. Exp. Psicol. Zumbido. Percepto. Realizar. 18, 1030–1044.
Resumen de Pubmed / Texto completo de Pubmed
Friedrich, F. J., Egly, R., Rafal, R. D., and Beck, D. (1998). Déficits de atención espacial en humanos: una comparación de lesiones superiores de unión parietal y temporal-parietal. Neuropsicología 12, 193-207.
Pubmed Abstract | Pubmed Texto Completo
Geng, J. J., y Behrmann, M. (2002). La detección de probabilidad de la ubicación del objetivo facilita implícitamente la búsqueda visual en participantes normales y pacientes con negligencia hemispacial. Psicol. sci. 13, 520–525.
Pubmed Abstract | Pubmed Texto Completo
Geng, J. J., y Behrmann, M. (2005). Probabilidad espacial como señal de atención en la búsqueda visual. Percepto. Psicófisis. 67, 1252–1268.
Resumen de Pubmed / Texto completo de Pubmed
Geng, J. J., and Mangun, G. R. (2011). La activación de la unión temporoparietal derecha mediante una señal contextual saliente facilita la discriminación del objetivo. Neuroimagen 54, 594-601.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Gibson, B. S., and Kelsey, E. M. (1998). La captura de atención impulsada por estímulos depende del conjunto de atención para las características visuales de toda la pantalla. J. Exp. Psicol. Zumbido. Percepto. Realizar. 24, 699–706.
Resumen de Pubmed / Texto completo de Pubmed
Giesbrecht, B., Woldorff, M. G., Song, A. W., and Mangun, G. R. (2003). Mecanismos neuronales de control de arriba hacia abajo durante la atención espacial y característica. Neuroimagen 19, 496-512.
Resumen de Pubmed / Texto completo de Pubmed / Texto completo cruzado
Gnadt, J. W., and Andersen, R. A. (1988). Actividad de planificación motora relacionada con la memoria en la corteza parietal posterior del macaco. Exp. Brain Res .. 70, 216-220.
Resumen de Pubmed / Texto completo de Pubmed
Gottlieb, J., and Balan, P. (2010). La atención como decisión en el espacio de información. Trends Cogn. Sci. 14, 240–248.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Greenberg, A. S., Esterman, M., Wilson, D., Serences, J. T. y Yantis, S. (2010). Control de la atención espacial y basada en características en la corteza frontoparietal. J. Neurocirujano. 30, 14330–14339.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Hopfinger, J. B., Buonocore, M. H., and Mangun, G. R. (2000). Los mecanismos neuronales del control atencional de arriba hacia abajo. NAT. Neurociencia. 3, 284–291.
Pubmed Abstract | Pubmed Texto Completo | CrossRef Texto Completo
Ipata, A. E., Gee, A. L., Gottlieb, J., Bisley, J. W., and Goldberg, M. E. (2006). Las respuestas de los labios a un estímulo emergente se reducen si se ignora abiertamente. NAT. Neurociencia. 9, 1071–1076.
Resumen de Pubmed / Texto completo de Pubmed / Texto completo cruzado
Johnston, W. A., and Dark, V. J. (1986). Atención Selectiva. Annu. Reverendo Psychol. 37, 43–75.
Juola, J. F., Koshino, H., and Warner, C. B. (1995). Compensaciones entre los efectos atencionales de las señales espaciales y los embates abruptos. Percepto. Psicófisis. 57, 333–342.
Resumen de Pubmed/Texto completo de Pubmed
Kastner, S., Pinsk, M. A., De Weerd, P., Desimone, R., y Ungerleider, L. G. (1999). Aumento de la actividad en la corteza visual humana durante la atención dirigida en ausencia de estimulación visual. Neurona 22, 751-761.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
Koechlin, E., Ody, C., and Kouneiher, F. (2003). La arquitectura del control cognitivo en la corteza prefrontal humana. Science 302, 1181-1185.
Pubmed Abstract | Pubmed Texto Completo | CrossRef Texto Completo
Koshino, H., Warner, C. B., and Juola, J. F. (1992). Efectividad relativa de señales centrales, periféricas y de inicio abrupto en la atención visual. Q. J. Exp. Psicol. A 45, 609-631.
Resumen de Pubmed/Texto completo de Pubmed
Liu, T., Slotnick, S. D., Serences, J. T., and Yantis, S. (2003). Mecanismos corticales de control atencional basado en características. Cereb. Cortex 13, 1334-1343.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Marois, R., Leung, H. C., and Gore, J.C. (2000). Un enfoque impulsado por estímulos para el procesamiento de la identidad y ubicación del objeto en el cerebro humano. Neurona 25, 717-728.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
McFire, J., and Zangwill, O. L. (1960). Discapacidades visuoconstructivas asociadas con lesiones del hemisferio cerebral izquierdo. Cerebro 82, 243-260.
Moore, C. M., and Egeth, H. (1998). ¿Cómo afecta la atención basada en funciones al procesamiento visual? J. Exp. Psicol. Zumbido. Percepto. Realizar. 24, 1296–1310.
Pubmed Abstract | Pubmed Texto Completo
Moran, J., and Desimone, R. (1985). La atención selectiva puertas de procesamiento visual en la corteza extraestriada. Science 229, 782-784.
Pubmed Abstract / Pubmed Full Text / CrossRef Full Text
O’Craven, K., Rosen, B. R., Kwong, K. K., Treisman, A., and Savoy, R. L. (1997). La atención voluntaria modula la actividad de la IRMF en la MT-MST humana. Neurona 18, 591-598.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Piercy, M. (1964). The effects of cerebral lesions on intellectual function: a review of current research trends (en inglés). Br. J. Psychiatry 110, 310-352.
Pubmed Resumen | Texto completo de Pubmed
Posner, M. I. (1980). Orientación de la atención. Q. J. Exp. Psicol. 32, 3–25.
Pubmed Abstract / Pubmed Full Text
Posner, M. I., Snyder, C. R., and Davidson, B. J. (1980). Atención y detección de señales. J. Exp. Psicol. 109, 160–174.
Pubmed Abstract / Pubmed Full Text
Posner, M. I., Walker, J. A., Friedrich, F. J., and Rafal, R. D. (1984). Efectos de la lesión parietal en la orientación encubierta de la atención. J. Neurocirujano. 4, 1863–1874.
Resumen de Pubmed / Texto completo de Pubmed
Serences, J. T., Shomstein, S., Leber, A. B., Golay, X., Egeth, H. E., and Yantis, S. (2005). Coordinación del control de atención voluntario e impulsado por estímulos en la corteza humana. Psicol. Sci. 16, 114–122.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Shaw, M. L. (1978). Un modelo de asignación de capacidad para el tiempo de reacción. J. Exp. Psicol. Zumbido. Percepto. Realizar. 4, 586–598.
Shomstein, S., y Behrmann, M. (2006). Sistemas corticales que median la atención visual a objetos y ubicaciones espaciales. Proc. Natl. Acad. Sci. U. S. A. 103, 11387-11392.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Shomstein, S., Lee, J., and Behrmann, M. (2010). Guía de atención de arriba hacia abajo y de abajo hacia arriba: investigando el papel de las cortezas parietales dorsal y ventral. Exp. Brain Res. 206, 197-208.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Shomstein, S. y Yantis, S. (2004a). Priorización configurable y contextual en atención basada en objetos. Psychon. Toro. Apo. 11, 247-253.
Resumen de Pubmed / Texto completo de Pubmed
Shomstein, S., and Yantis, S. (2004b). El control de la atención cambia entre la visión y la audición en la corteza humana. J. Neurocirujano. 24, 10702–10706.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Shomstein, S., y Yantis, S. (2006). La corteza parietal media el control voluntario de la atención auditiva espacial y no espacial. J. Neurocirujano. 26, 435–439.
Resumen de Pubmed / Texto Completo de Pubmed / Texto cruzado
Silver, M. A., Ress, D., and Heeger, D. J. (2005). Mapas topográficos de la atención espacial visual en la corteza parietal humana. J. Neurofisiol. 94, 1358–1371.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Tamber-Rosenau, B. J., Esterman, M., Chiu, Y.-C., y Yantis, S. (2011). Mecanismos corticales de control cognitivo para cambiar la atención en la visión y la memoria de trabajo. J. Cogn. Neurociencia. 23, 2905–2919.
Resumen de Pubmed / Texto Completo de Pubmed / Texto completo cruzado
Theeuwes, J. (1991). Control exógeno y endógeno de la atención: el efecto de on y compensaciones visuales. Percepto. Psicófisis. 49, 83–90.
Resumen de Pubmed / Texto completo de Pubmed
Vallar, G., and Perani, D. (1986). Anatomía de la negligencia unilateral después de lesiones por accidente cerebrovascular del hemisferio derecho. Estudio de correlación clínico / tomografía computarizada en hombre. Neuropsicología 24, 609-622.
Resumen de Pubmed / Texto completo de Pubmed / Texto completo cruzado
Wojciulik, E., and Kanwisher, N. (1999). La generalidad de la participación parietal en la atención visual. Neurona 23, 747-764.
Resumen de Pubmed / Texto completo de Pubmed/Texto completo cruzado
Yantis, S., and Jonides, J. (1984). Escenarios visuales abruptos y atención selectiva: evidencia de la búsqueda visual. J. Exp. Psicol. Zumbido. Percepto. Realizar. 10, 601–621.
Resumen de Pubmed / Texto completo de Pubmed
Yantis, S., and Jonides, J. (1990). Escenarios visuales abruptos y atención selectiva: asignación voluntaria versus automática. J. Exp. Psicol. Zumbido. Percepto. Realizar. 16, 121–134.
Pubmed Abstract | Pubmed Texto Completo
Yantis, S., Schwarzbach, J., Serences, J. T., Carlson, R. L., Steinmetz, M. A., Pekar, J. J., and Courtney, S. M. (2002). Actividad neural transitoria en la corteza parietal humana durante los cambios de atención espacial. NAT. Neurociencia. 5, 995–1002.
Resumen de Pubmed / Texto Completo de Pubmed/Texto completo cruzado
Yantis, S., y Serences, J. T. (2003). Mecanismos corticales de control de la atención basada en el espacio y en objetos. Curr. Opin. Neurobiol. 13, 187–193.
Resumen de Pubmed / Texto Completo de Pubmed / Texto Cruzado