selvom Rubisco er ansvarlig for langt størstedelen af organisk kulstof på jordens overflade, kan dets iltaseaktivitet alvorligt reducere fotosyntetisk effektivitet. Nogle planter har udviklet en måde at minimere RuBisCOs iltaseaktivitet på.
læringsmål
- Identificer de forhold, der øger RuBisCO ‘s iltegenaseaktivitet
- Beskriv, hvordan RuBisCO’ s iltegenaseaktivitet reducerer fotosyntetisk effektivitet
- skelne mellem C3-og C4-ordninger til kulstoffiksering
- Afvej fordele og ulemper ved C3 versus C4
- Sammenlign og kontrast fotosyntese og respiration og deres forhold i de globale kulstof-og iltcyklusser.
RuBisCOs iltase-aktivitet forringer fotosyntetisk effektivitet
Rubisco har iltase-aktivitet såvel som carboksylaseaktivitet; det løser undertiden O2 til RuBP i stedet for CO2. Iltaseaktiviteten forekommer ved lave CO2, høje O2-forhold og bliver udtalt ved høje temperaturer. Som et resultat iltes organisk kulstof, det modsatte af fotosyntese, hvilket reducerer uorganisk kulstof for at fremstille organisk kulstof.
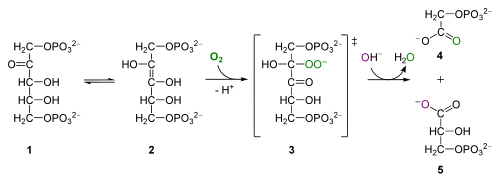
for de nysgerrige: iltase aktivitet af Rubisco, fra
iltning af RuBP producerer 2-phosphoglycolat, en 2-carbon toksisk forbindelse, der gennemgår en række reaktioner i peroksisomet og mitokondrier, frigiver CO2 og resulterer i tab af organisk kulstof og energiproduktion. Denne proces kaldes fotorespiration – et forfærdeligt vildledende navn for studerende, fordi det ikke har noget at gøre med åndedræt og giver ingen ATP. Alle Biol 1510-studerende skal huske om fotorespiration er, at det reducerer fotosyntetisk effektivitet, og at det opstår, når Rubisco ilter RuBP i stedet for carboksylerende RuBP.
Rubisco udviklede sig allerede før ilt fotosyntese, da der ikke var ilt i atmosfæren eller i havets farvande, så der var ingen udvælgelse mod iltaseaktivitet. Ikke desto mindre har hverken natur eller human genteknologi i over 2 milliarder år været i stand til at eliminere eller endda reducere Rubisco ‘ s iltaseaktivitet markant uden også at påvirke carboksylaseaktiviteten.
C4-planter har udviklet en mekanisme til at levere CO2 til Rubisco
for at planter kan optage CO2, skal de åbne strukturer kaldet stomata på deres blade, som er porer, der tillader gasudveksling. Planter mister også vanddamp gennem deres stomata, hvilket betyder, at de kan dø af dehydrering under tørre forhold, da de holder deres stomata åben for fotosyntese. Som reaktion lukker planter deres stomata for at forhindre dehydrering. De umiddelbare * konsekvenser af lukket stomata på kort sigt er reduceret CO2-koncentration (CO2-niveauer falder, når det bruges til fotosyntese) og øget O2-koncentration (O2-niveauer stiger, når det produceres ved fotosyntese). De stigende O2-niveauer øger hastigheden af fotorespiration (reaktion af rubisco med ilt i stedet for kulsyre), når derefter drastisk reducerer effektiviteten af rubisco, som allerede er et meget langsomt fungerende middel. Så det betyder, at planter under tørre forhold risikerer dehydrering, hvis de åbner deres stomata for at fremme gasudveksling eller manglende evne til at producere sukker, hvis de holder deres stomata lukket for at minimere dehydrering. (*den langsigtede konsekvens af permanent lukket stomata er død ved kvælning, da mitokondrier løber tør for ilt for at udføre respiration.)
mange planter, der lever under varme og/eller tørre forhold, har udviklet en alternativ kulstoffikseringsvej for at forbedre effektiviteten af rubisco, så de ikke behøver at holde deres stomata åbne så meget, og dermed reducerer de risikoen for at dø af dehydrering. Disse planter kaldes C4-planter, fordi det første produkt af kulstoffiksering er en 4-carbonforbindelse (i stedet for en 3-carbonforbindelse som i C3 eller “normale” planter). C4 planter bruger denne 4-carbon forbindelse til effektivt at” koncentrere ” CO2 omkring rubisco, så rubisco er mindre sandsynligt re reagere med O2.
Der er to vigtige tilpasninger, der gør det muligt for C4-planter at gøre dette:
- for det første bruger C4-planter et alternativt ferment til det første trin i kulstoffiksering. Det har ingen iltaktivitet og har en meget højere affinitet for CO2 end rubisco. Som navnet “PEP carboksylase” antyder, binder det CO2 til en forbindelse kaldet phosphoenolpyruvat (PEP).
- for det andet har C4-planter specialiseret bladanatomi med to forskellige typer fotosyntetiske celler: mesophyll celler (på ydersiden af bladet, nær stomata) og bundt kappe celler (i det indre af bladet, langt væk fra stomata). Rubisco er placeret i bundtkappeceller, men ikke i mesofylceller.
Sådan fungerer alt dette:
PEP carboksylase er placeret i mesofylcellerne på bladets ydre nær stomata. Der er ingen rubisco i mesofylcellerne. CO2, der kommer ind i stomata, fastgøres hurtigt af PEP carboksylase i en 4-carbonforbindelse, kaldet malat, ved at fastgøre CO2 til PEP. Malatet transporteres derefter dybere ind i bladvævet til bundtkappecellerne, som begge er langt væk fra stomata (og dermed langt væk fra ilt) og indeholder rubisco. En gang inde i bundtkappecellerne dekarboksyleres malat for at frigive pyruvat og CO2; CO2 fastgøres derefter af rubisco som en del af Calvin-cyklussen, ligesom i C3-planter. Pyruvat vender derefter tilbage til mesofylcellerne, hvor et fosfat fra ATP bruges til at regenerere PEP. Således i C4-anlæg, C4 kulstoffiksering har en nettotilvækst på 1 ATP for hver CO2 leveret til rubisco; Imidlertid er C4-planter mindre tilbøjelige til at dø af dehydrering sammenlignet med C3-planter under tørre forhold.
billedet nedenfor illustrerer den vej, vi netop har beskrevet:
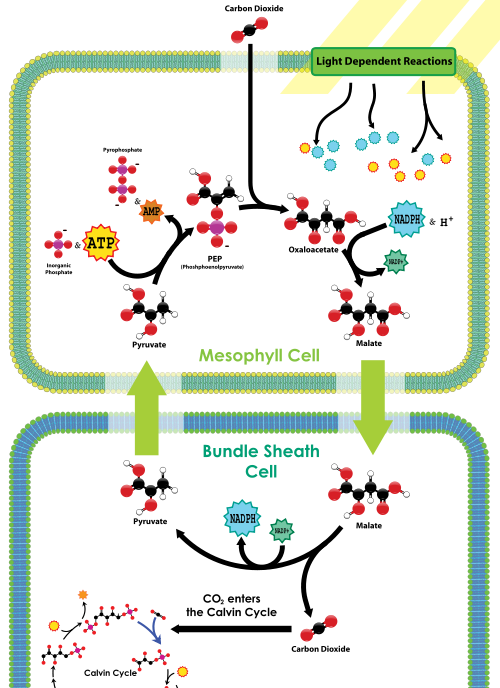
C4 carbonfikseringsoversigt, fra
hvad Biol 1510-studerende skal huske om C4 er, at disse planter har tilføjet en CO2-koncentrationsmekanisme til at fodre rubisco og Calvin-cyklussen; mekanismen bruger pep carboksylase til oprindeligt at fremstille en 4-carbonforbindelse, der derefter frigiver CO2 til RuBisCO i bladceller, der udsættes for lidt ilt. Mens denne mekanisme reducerer RuBisCOs iltaseaktivitet, har den en ekstra energikostnad i form af en anden ATP pr.
den (beskårne) video nedenfor giver en stor oversigt sammenligning af C3 vs C4 fotosyntese, men vær opmærksom på følgende problemer med denne video: videoen får det til at lyde som om RuBP katalyserer sin egen reaktion med CO2 til at danne til 2 molekyler af 3-carbon 3PG, i stedet for at være en af reaktanterne i denne reaktion. Denne reaktion katalyseres af rubisco, og det er rubisco, der har iltningsaktivitet, ikke RuBP! Videoen får det også til at lyde som om PEP katalyserer sin egen reaktion med CO2 til dannelse af 4-carbonmalat, når denne reaktion i virkeligheden katalyseres af PEP carboksylase.
https://www.youtube.com/watch?v=HbLg4lMpUa
Hvis du vil vide mere, giver videoen nedenfor en mere grundig (omend lidt langsom) illustration af denne proces:
fordele og ulemper ved C4 – og C3-carbonfiksering
- C4-planter vokser bedre end C3-planter under varme, tørre forhold, når planter skal lukke deres stomata for at spare vand-med stomata lukket, CO2-niveauer i det indre af bladet falder, og O2-niveauer stiger.
- C3-planter vokser bedre end C4-planter under kølige, fugtige forhold, når planter kan åbne deres stomata, fordi C3-planter ikke pådrager sig ekstraomkostningerne ved ATP for C4-kulstoffiksering.
fotosyntese og Respiration: spejlbilleder
de kemiske ligninger for ilt fotosyntese og aerob respiration er nøjagtigt det modsatte af hinanden.
en balance mellem de globale fotosyntesehastigheder (primærproduktion) og globale respirationshastigheder er nødvendig for at opretholde stabile atmosfæriske koncentrationer af CO2 og O2.i eukaryoter forekommer både fotosyntese og respiration i organeller med dobbeltmembraner og deres egne cirkulære genomer, der stammer fra prokaryote endosymbionter.
begge processer har elektrontransportkæder, kemiosmose og ATP-syntase drevet af protonmotivkraft.
de effektpoint-dias, der bruges i video-screencasts, er i carbonfikseringsdiassættet.