Även om Rubisco är ansvarig för den stora delen av organiskt kol på jordens yta, kan dess oxygenasaktivitet kraftigt minska fotosyntetisk effektivitet. Vissa växter har utvecklat ett sätt att minimera oxygenasaktiviteten hos Rubisco.
inlärningsmål
- identifiera de förhållanden som ökar oxygenasaktiviteten hos Rubisco
- Beskriv hur oxygenasaktiviteten hos Rubisco minskar fotosyntetisk effektivitet
- skilja C3-och C4-scheman för kolfixering
- väga fördelarna och nackdelarna med C3 kontra C4
- Jämför och kontrast fotosyntes och andning, och deras förhållande i de globala kol-och syrecyklerna.
Rubiscos oxygenasaktivitet försämrar fotosyntetisk effektivitet
Rubisco har oxygenasaktivitet såväl som karboxylasaktivitet; det fixar ibland O2 till RuBP istället för CO2. Oxygenasaktiviteten uppträder vid låga CO2, höga O2-förhållanden och uttalas vid höga temperaturer. Som ett resultat oxideras organiskt kol, motsatsen till fotosyntes, vilket minskar oorganiskt kol för att göra organiskt kol.
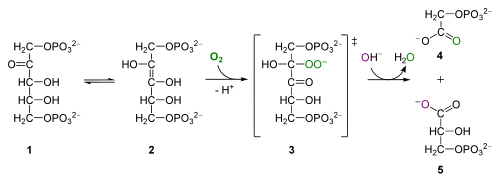
för nyfiken: oxygenasaktivitet av Rubisco, från Wikipedia. RuBP (1) interkonverterar till en Enol isomer (2) som kombinerar med syre för att bilda den instabila mellanprodukten (3) som hydrolyserar till fosfoglykolat (4) och 3PG (5)
syresättningen av RuBP producerar 2-fosfoglykolat, en 2-kol giftig förening som genomgår en serie reaktioner i peroxisomen och mitokondrier, frigör CO2 och resulterar i förlust av organiskt kol och energiproduktion. Denna process kallas photorespiration – ett väldigt vilseledande namn för studenter, eftersom det inte har något att göra med andning och ger ingen ATP. Alla Biol 1510-studenter behöver komma ihåg om fotorespiration är att det minskar fotosyntetisk effektivitet och att det uppstår när Rubisco oxygenerar RuBP istället för att karboxylera RuBP.
Rubisco utvecklades redan före oxygenisk fotosyntes, när det inte fanns något syre i atmosfären eller i havsvattnet, så det fanns inget val mot oxygenasaktivitet. Men på över 2 miljarder år har varken natur eller mänsklig genteknik kunnat eliminera eller till och med avsevärt minska oxygenasaktiviteten hos Rubisco utan att också påverka karboxylasaktiviteten.
C4-växter har utvecklat en mekanism för att leverera CO2 till Rubisco
för att växter ska ta in CO2 måste de öppna strukturer som kallas stomata på sina löv, vilka är porer som tillåter gasutbyte. Växter förlorar också vattenånga genom sin stomata, vilket innebär att de kan dö av uttorkning under torra förhållanden eftersom de håller sin stomata öppen för fotosyntes. Som svar stänger växterna sin stomata för att förhindra uttorkning. De omedelbara * konsekvenserna av sluten stomata på kort sikt är minskad CO2-koncentration (CO2-nivåer minskar när den används för fotosyntes) och ökad O2-koncentration (O2-nivåer ökar när den produceras genom fotosyntes). De stigande O2-nivåerna ökar graden av fotorespiration (reaktion av rubisco med syre istället för koldioxid), då drastiskt minskar effektiviteten hos rubisco, som redan är ett mycket långsamt arbetande enzym. Så det betyder att växter i torra förhållanden riskerar uttorkning om de öppnar sin stomata för att främja gasutbyte eller oförmåga att producera socker om de håller stomata stängda för att minimera uttorkning. (*den långsiktiga konsekvensen av permanent stängd stomata är död genom kvävning eftersom mitokondrierna tar slut på syre för att utföra andning.)
många växter som lever i varma och / eller torra förhållanden har utvecklat en alternativ kolfixeringsväg för att förbättra effektiviteten hos rubisco så att de inte behöver hålla stomata öppna så mycket, och därmed minskar risken för att dö av uttorkning. Dessa växter kallas C4-växter, eftersom den första produkten av kolfixering är en 4-kolförening (istället för en 3-kolförening som i C3 eller ”normala” växter). C4-växter använder denna 4-kolförening för att effektivt ”koncentrera” CO2 runt rubisco, så att rubisco är mindre sannolikt att reagera med O2.
det finns två viktiga anpassningar som gör det möjligt för C4-växter att göra detta:
- först använder C4-växter ett alternativt enzym för det första steget av kolfixering. Detta enzym kallas fosfoenolpyruvat (PEP) karboxylas, och det har ingen oxygenasaktivitet och har en mycket högre affinitet för CO2 än rubisco. Som namnet ”PEP-karboxylas” antyder fäster enzymet CO2 till en förening som kallas fosfoenolpyruvat (PEP).
- för det andra har C4-växter specialiserad bladanatomi med två olika typer av fotosyntetiska celler: mesofyllceller (på bladets utsida, nära stomata) och buntmantelceller (i bladets inre, långt borta från stomata). Rubisco ligger i buntmantelceller, men inte i mesofyllceller.
Så här fungerar allt detta:
PEP-karboxylas ligger i mesofyllcellerna, på bladets yttre nära stomata. Det finns ingen rubisco i mesofyllcellerna. CO2 som kommer in i stomata fixeras snabbt av PEP-karboxylas i en 4-kolförening, kallad malat, genom att fästa CO2 till PEP. Malatet transporteras sedan djupare in i bladvävnaden till buntmantelcellerna, som båda är långt borta från stomata (och därmed långt borta från syre) och innehåller rubisco. En gång inuti buntmantelcellerna dekarboxyleras malat för att frigöra pyruvat och CO2; CO2 fixeras sedan av rubisco som en del av Calvin-cykeln, precis som i C3-växter. Pyruvat återvänder sedan till mesofyllcellerna, där ett fosfat från ATP används för att regenerera PEP. Således i C4-växter har C4-kolfixering en nettokostnad på 1 ATP för varje CO2 som levereras till rubisco; C4-växter är dock mindre benägna att dö av uttorkning jämfört med C3-växter under torra förhållanden.
bilden nedan illustrerar vägen vi just beskrivit:
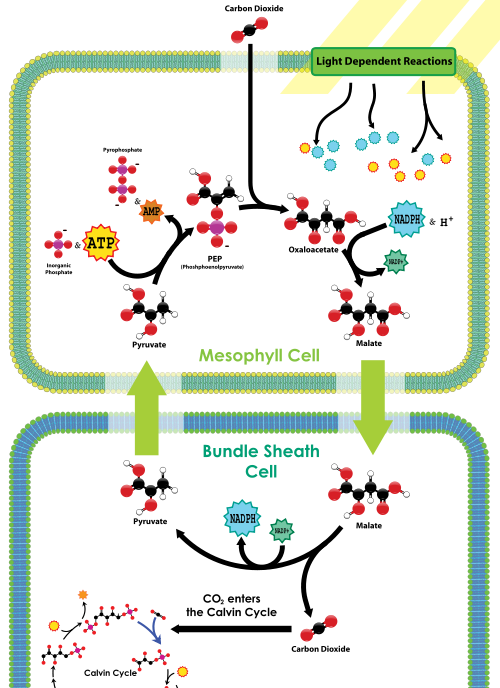
C4 Carbon fixation översikt, från Wikimedia
vad Biol 1510-studenter behöver komma ihåg om C4 är att dessa växter har lagt till en CO2-koncentrationsmekanism för att mata rubisco och Calvin-cykeln; mekanismen använder Pep-karboxylas för att initialt göra en 4-kolförening, som sedan släpper ut CO2 till rubisco i bladceller som utsätts för lite syre. Medan denna mekanism minskar oxygenasaktiviteten hos rubisco, har den en extra energikostnad i form av en annan ATP per mol CO2 fixerad.
den (beskurna) videon nedan ger en bra översiktsjämförelse av C3 vs C4 fotosyntes, men var medveten om följande problem med den här videon: videon gör att det låter som om RuBP katalyserar sin egen reaktion med CO2 för att bilda till 2 molekyler av 3-kol 3PG, istället för att vara en av reaktanterna i denna reaktion. Denna enzymatiska reaktion katalyseras av rubisco, och det är rubisco som har oxygnasaktivitet, inte RuBP! Videon gör också att det låter som att PEP katalyserar sin egen reaktion med CO2 för att bilda 4-kolmalat, när denna reaktion i verkligheten katalyseras av PEP-karboxylas.
https://www.youtube.com/watch?v=HbLg4lMpUa
om du vill veta mer ger videon nedan en mer grundlig (om än något långsam) illustration av denna process:
fördelar och nackdelar med C4 – och C3-kolfixering
- C4-växter växer bättre än C3-växter under heta, torra förhållanden när växter måste stänga sin stomata för att spara vatten-med stomata stängd, CO2-nivåer i bladets inre fall och O2-nivåer stiger.
- C3-växter växer bättre än C4-växter under svala, fuktiga förhållanden när växter kan öppna sin stomata, eftersom C3-växter inte medför extra kostnader för ATP för C4-kolfixering.
fotosyntes och andning: spegelbilder
de kemiska ekvationerna för Oxygen fotosyntes och aerob andning är exakt motsatsen till varandra.
en balans mellan de globala fotosynteshastigheterna (primärproduktion) och globala andningshastigheter behövs för att upprätthålla stabila atmosfäriska koncentrationer av CO2 och O2.
i eukaryoter förekommer både fotosyntes och andning i organeller med dubbla membran och deras egna cirkulära genom, som har sitt ursprung som prokaryota endosymbionter.
båda processerna har elektrontransportkedjor, kemiosmos och ATP-syntas som drivs av protonmotivkraft.
powerpoint-bilderna som används i videoskärmsläpparna finns i bilduppsättningen för kolfixering.