chociaż Rubisco jest odpowiedzialny za ogromną większość węgla organicznego na powierzchni Ziemi, jego aktywność tlenazy może poważnie zmniejszyć wydajność fotosyntetyczną. Niektóre rośliny wyewoluowały sposób na zminimalizowanie aktywności oksygenazy Rubisco.
cele nauki
- Zidentyfikuj warunki, które zwiększają aktywność oksygenazy Rubisco
- opisz, w jaki sposób aktywność oksygenazy Rubisco zmniejsza wydajność fotosyntezy
- rozróżnij Schematy C3 I C4 dla utrwalenia węgla
- zważaj zalety i wady C3 w porównaniu z C4
- Porównaj i kontrastuj fotosyntezę i oddychanie oraz ich związek w globalnych cyklach węgla i tlenu.
aktywność oksygenazy Rubisco pogarsza wydajność fotosyntetyczną
Rubisco ma aktywność oksygenazy, a także karboksylazy; czasami naprawia O2 do RuBP zamiast CO2. Aktywność oksygenazy występuje w warunkach niskiego CO2, Wysokiego O2 i staje się wyraźna w wysokich temperaturach. W rezultacie węgiel organiczny jest utleniany, co jest przeciwieństwem fotosyntezy, która redukuje węgiel nieorganiczny do węgla organicznego.
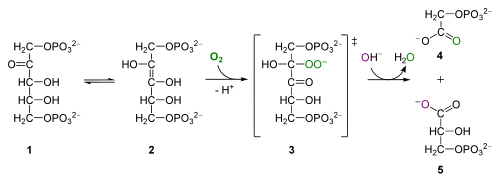
dla ciekawych: aktywność oksygenazy Rubisco, z Wikipedii. RuBP (1) interkonwertuje do izomeru enolu (2), który łączy się z tlenem, tworząc niestabilny związek pośredni (3), który hydrolizuje do fosfoglikolanu (4) i 3PG (5)
natlenianie RuBP wytwarza 2-fosfoglikolan, toksyczny związek 2-węglowy, który przechodzi szereg reakcji w peroksysomie i mitochondriach, uwalniając CO2 i powodując utratę węgla organicznego i produkcję energii. Proces ten nazywany jest fotorespiracją – strasznie myląca nazwa dla studentów, ponieważ nie ma on nic wspólnego z oddychaniem i nie daje ATP. Wszyscy uczniowie Biol 1510 powinni pamiętać o tym, że fotorespiracja zmniejsza efektywność fotosyntetyczną i że występuje, gdy Rubisco dotlenia RuBP zamiast Karboksylowania RuBP.
Rubisco wyewoluował jeszcze przed fotosyntezą tlenową, kiedy nie było tlenu w atmosferze ani w wodach oceanicznych, więc nie było selekcji przeciwko aktywności oksygenazy. Niemniej jednak w ciągu ponad 2 miliardów lat ani natura, ani ludzka inżynieria genetyczna nie były w stanie wyeliminować lub nawet znacznie zmniejszyć aktywności oksygenazy Rubisco bez wpływu na aktywność karboksylazy.
rośliny C4 wyewoluowały mechanizm dostarczania CO2 do Rubisco
aby rośliny mogły przyjmować CO2, muszą otwierać na liściach struktury zwane szparkami, które są porami umożliwiającymi wymianę gazową. Rośliny tracą również parę wodną przez aparaty szparkowe, co oznacza, że mogą umrzeć z powodu odwodnienia w suchych warunkach, ponieważ utrzymują aparaty szparkowe otwarte na fotosyntezę. W odpowiedzi rośliny zamykają aparaty szparkowe, aby zapobiec odwodnieniu. Natychmiastowe * konsekwencje zamkniętych aparatów szparkowych w krótkim okresie są zmniejszone stężenie CO2 (poziom CO2 zmniejsza się, gdy jest wykorzystywany do fotosyntezy) i zwiększone stężenie O2 (poziom O2 wzrasta, gdy jest wytwarzany przez fotosyntezę). Rosnący poziom O2 zwiększa szybkość fotorespiracji (reakcja rubisco z tlenem zamiast dwutlenku węgla), kiedy to drastycznie zmniejsza wydajność rubisco, który jest już bardzo wolno działającym enzymem. Oznacza to, że rośliny w suchych warunkach są narażone na odwodnienie, jeśli otwierają aparaty szparkowe, aby promować wymianę gazową, lub niezdolność do produkcji cukru, jeśli trzymają aparaty szparkowe zamknięte, aby zminimalizować odwodnienie. (*długotrwałą konsekwencją trwale zamkniętego aparatu szparkowego jest śmierć przez uduszenie, gdy mitochondriom zabraknie tlenu do przeprowadzenia oddychania.)
wiele roślin, które żyją w gorących i / lub suchych warunkach, rozwinęło alternatywną ścieżkę wiązania węgla, aby zwiększyć wydajność produktu rubisco, dzięki czemu nie muszą utrzymywać swoich aparatów szparkowych tak bardzo otwartych, a tym samym zmniejszają ryzyko śmierci z powodu odwodnienia. Rośliny te nazywane są roślinami C4, ponieważ pierwszym produktem wiązania węgla jest związek 4-węglowy (zamiast związku 3-węglowego jak w roślinach C3 lub „normalnych”). Rośliny C4 używają tego 4-węglowego związku, aby skutecznie „skoncentrować” CO2 wokół rubisco, dzięki czemu rubisco rzadziej reaguje z O2.
istnieją dwie ważne adaptacje, które pozwalają roślinom C4 to zrobić:
- po pierwsze, rośliny C4 używają alternatywnego enzymu do pierwszego etapu wiązania węgla. Enzym ten nazywa się karboksylazą fosfoenolopirogronianową (PEP) i nie ma aktywności oksygenazy i ma znacznie większe powinowactwo do CO2 niż rubisco. Jak sugeruje nazwa” karboksylaza PEP”, enzym przyłącza CO2 do związku zwanego fosfoenolopirogronianem (PEP).
- Po drugie, rośliny C4 mają wyspecjalizowaną anatomię liści z dwoma różnymi typami komórek fotosyntetycznych: komórki mezofilne (Na Zewnątrz liścia, w pobliżu aparatów szparkowych) i komórki osłonkowe (we wnętrzu liścia, z dala od aparatów szparkowych). Rubisco znajduje się w komórkach osłonek pęczkowych, ale nie w komórkach mezofilnych.
oto jak to wszystko działa:
karboksylaza PEP znajduje się w komórkach mezofilnych, na zewnętrznej stronie liścia, w pobliżu aparatów szparkowych. W komórkach mezofilu nie ma rubisco. CO2 wchodzący do aparatów szparkowych jest szybko utrwalany przez karboksylazę PEP do Związku 4-węglowego, zwanego jabłczanem, poprzez przyłączenie CO2 do PEP. Jabłczan jest następnie transportowany głębiej w tkankę liściową do komórek osłonki pęczka, które są zarówno daleko od aparatów szparkowych (a więc daleko od tlenu) i zawierają rubisco. Po wejściu do komórek powłoki wiązki jabłczan jest dekarboksylowany w celu uwolnienia pirogronianu i CO2; CO2 jest następnie ustalany przez rubisco w ramach cyklu Calvina, podobnie jak u roślin C3. Następnie pirogronian wraca do komórek mezofilu, gdzie fosforan z ATP jest używany do regeneracji PEP. Tak więc w instalacjach C4 Wiązanie węgla C4 ma dodatkowy koszt netto w wysokości 1 ATP za każdy CO2 dostarczony do rubisco; jednak rośliny C4 są mniej narażone na śmierć z powodu odwodnienia w porównaniu do roślin C3 w suchych warunkach.
poniższy obraz ilustruje ścieżkę, którą właśnie opisaliśmy:
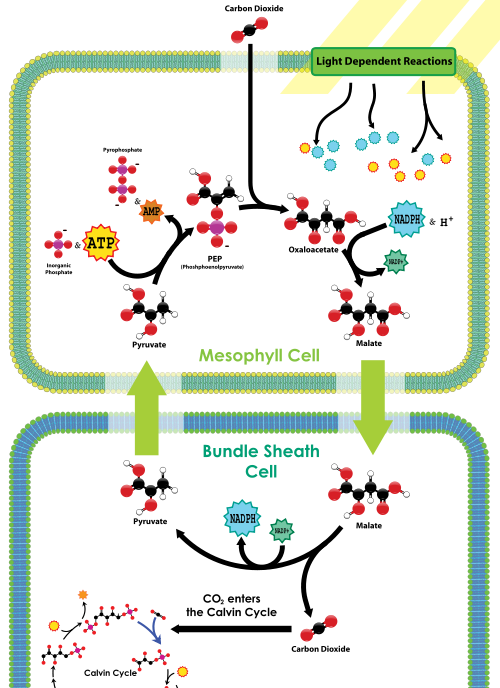
przegląd wiązania węgla C4 z Wikimedia
uczniowie Biol 1510 muszą pamiętać o C4, że te rośliny dodały mechanizm koncentracji CO2 do zasilania rubisco i cyklu Calvina; mechanizm wykorzystuje karboksylazę pep, aby początkowo wytworzyć związek 4-węglowy, który następnie uwalnia CO2 do rubisco w komórkach liści, które są narażone na niewielką ilość tlenu. Chociaż mechanizm ten zmniejsza aktywność oksygenazy rubisco, ma on dodatkowy koszt energii w postaci innego ATP na mol CO2.
poniższy (przycięty) film przedstawia świetne porównanie fotosyntezy C3 I C4, ale należy pamiętać o następujących problemach związanych z tym filmem: film sprawia, że brzmi on tak, jakby RuBP katalizował własną reakcję z CO2, tworząc 2 cząsteczki 3-węgla 3PG, zamiast być jednym z reagentów w tej reakcji. Ta reakcja enzymatyczna jest katalizowana przez rubisco i to rubisco ma aktywność oksygnazy, a nie RuBP! Wideo sprawia również, że brzmi to tak, jakby PEP katalizuje własną reakcję z CO2, tworząc 4-węglowy jabłczan, podczas gdy w rzeczywistości reakcja ta jest katalizowana przez karboksylazę PEP.
https://www.youtube.com/watch?v=HbLg4lMpUa
Jeśli chcesz dowiedzieć się więcej, poniższy film przedstawia dokładniejszą (choć nieco powolną) ilustrację tego procesu:
zalety i wady wiązania węgla C4 i C3
- rośliny C4 rosną lepiej niż rośliny C3 w gorących, suchych warunkach, gdy rośliny muszą zamknąć aparaty szparkowe, aby oszczędzać wodę – przy zamkniętych aparatach szparkowych poziom CO2 we wnętrzu liści spada, a poziom O2 wzrasta.
- rośliny C3 rosną lepiej niż rośliny C4 w chłodnych, wilgotnych warunkach, gdy rośliny mogą otworzyć swoje aparaty szparkowe, ponieważ rośliny C3 nie ponoszą dodatkowych kosztów ATP wiązania węgla C4.
fotosynteza i oddychanie: odbicia lustrzane
równania chemiczne fotosyntezy tlenowej i oddychania tlenowego są dokładnie odwrotne do siebie.
równowaga między globalnym tempem fotosyntezy (produkcja pierwotna) a globalnym tempem oddychania jest potrzebna do utrzymania stabilnego stężenia CO2 i O2 w atmosferze.
u eukariotów zarówno fotosynteza, jak i oddychanie występują w organelach z podwójnymi błonami i ich własnymi kolistymi genomami, które powstały jako prokariotyczne endosymbionty.
oba procesy mają łańcuchy transportu elektronów, chemiosmozę i syntazę ATP napędzaną siłą napędową protonów.
slajdy powerpoint używane w screencastach wideo są w zestawie ślizgowym do mocowania Węglowego.