vaikka Rubisco on vastuussa valtavasta osasta maan pinnalla olevaa orgaanista hiiltä, sen oksigenaasiaktiivisuus voi merkittävästi heikentää fotosynteettistä tehokkuutta. Jotkin kasvit ovat kehittäneet tavan minimoida rubiscon oksigenaasiaktiivisuus.
oppimistavoitteet
- tunnista rubiscon oksigenaasiaktiivisuutta lisäävät olosuhteet
- kuvaavat, miten rubiscon oksigenaasiaktiivisuus vähentää fotosynteettistä tehokkuutta
- erottaa C3-ja C4-järjestelmät hiilen sitomiseksi
- punnitsevat C3: n ja C4: n etuja ja haittoja
- vertaa ja vertaile fotosynteesiä ja hengitystä sekä niiden suhdetta maailmanlaajuisissa hiili-ja happisykleissä.
rubiscon oksigenaasiaktiivisuus heikentää fotosynteettistä tehokkuutta
rubiscon oksigenaasiaktiivisuus samoin kuin karboksylaasiaktiivisuus; joskus se korjaa O2: n RuBP: ksi CO2: n sijaan. Oksigenaasiaktiivisuus tapahtuu matalissa CO2-ja korkeissa O2-olosuhteissa ja korostuu korkeissa lämpötiloissa. Tämän seurauksena Orgaaninen hiili hapettuu, mikä on fotosynteesin vastakohta, jolloin epäorgaaninen hiili pelkistyy orgaaniseksi hiileksi.
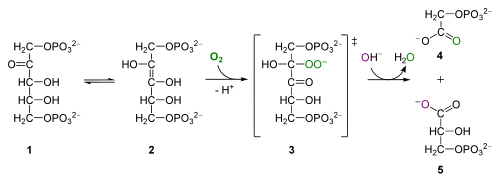
for the curious: oxygenase activity of Rubisco, Wikipediasta. RuBP (1) transkonvertoituu enoli-isomeeriksi (2), joka yhdistyy hapen kanssa muodostaen epästabiilin välituotteen (3), joka hydrolysoituu fosfoglykolaatiksi (4) ja 3PG (5)
rubp: n hapetus tuottaa 2-fosfoglykolaattia, 2-hiilistä myrkyllistä yhdistettä, joka käy läpi sarjan reaktioita peroksisomissa ja mitokondrioissa vapauttaen CO2: ta ja johtaen orgaanisen hiilen ja energiantuotannon menetykseen. Tätä prosessia kutsutaan fotorespiraatioksi-hirveän harhaanjohtava nimi opiskelijoille, koska sillä ei ole mitään tekemistä hengityksen kanssa eikä se tuota ATP: tä. Kaikki Biol 1510 opiskelijoiden täytyy muistaa fotorespiraatio on, että se vähentää fotosynteettistä tehokkuutta, ja että se tapahtuu, kun Rubisco hapettaa RuBP sijaan karboksyloimalla RuBP.
Rubisco kehittyi jo ennen oksigeenistä fotosynteesiä, jolloin ilmakehässä tai merivesissä ei ollut happea, joten oksigenaasiaktiivisuutta vastaan ei ollut valintaa. Yli 2 miljardissa vuodessa luonto tai ihmisen geenitekniikka ei kuitenkaan ole kyennyt poistamaan tai edes merkittävästi vähentämään rubiscon oksigenaasiaktiivisuutta vaikuttamatta myös karboksylaasiaktiivisuuteen.
C4-kasvit ovat kehittäneet mekanismin toimittaa CO2 Rubiscoon
, jotta kasvit voisivat ottaa CO2: ta, niiden on avattava lehdissään ilmarakenteita, jotka ovat kaasunvaihdon mahdollistavia huokosia. Kasvit menettävät myös vesihöyryä ilmansa kautta, mikä tarkoittaa sitä, että ne voivat kuivissa olosuhteissa kuolla nestehukkaan, kun ne pitävät ilmansa auki fotosynteesiä varten. Vastauksena kasvit sulkevat ilmansa kuivumisen estämiseksi. Suljetun ilmatilan välittömät * seuraukset lyhyellä aikavälillä ovat vähentynyt CO2-pitoisuus (CO2-tasot vähenevät, kun sitä käytetään fotosynteesissä) ja lisääntynyt O2-pitoisuus (O2-tasot kasvavat, kun sitä tuotetaan fotosynteesissä). Kohoavat O2-pitoisuudet lisäävät fotorespiraationopeutta (rubiscon reaktio hapen kanssa hiilidioksidin sijaan), jolloin rubiscon, joka on jo hyvin hitaasti toimiva entsyymi, tehokkuus heikkenee rajusti. Tämä tarkoittaa siis sitä, että kuivissa olosuhteissa elävät kasvit ovat vaarassa kuivua, jos ne avaavat ilmansa edistääkseen kaasun vaihtoa, tai kyvyttömiä tuottamaan sokeria, jos ne pitävät ilmansa suljettuna nestehukan minimoimiseksi. (*pysyvästi suljetun ilmatilan pitkäaikainen seuraus on tukehtumiskuolema, kun mitokondrioista loppuu happi hengityksen suorittamiseksi.)
monet kuumissa ja / tai kuivissa olosuhteissa elävät kasvit ovat kehittäneet vaihtoehtoisen hiilensidontareitin rubiscon tehokkuuden parantamiseksi, jotta niiden ei tarvitse pitää vatsaansa yhtä paljon auki, ja näin ne vähentävät riskiä kuolla nestehukkaan. Näitä kasveja kutsutaan C4-kasveiksi, koska hiilensidonnan ensimmäinen tuote on 4-Hiilinen yhdiste (eikä 3-Hiilinen yhdiste kuten C3: ssa tai ”normaaleissa” kasveissa). C4-kasvit käyttävät tätä 4-hiilistä yhdistettä tehokkaasti” konsentroimaan ” CO2: ta rubiscon ympärille, joten rubisco ei todennäköisesti reagoi uudelleen O2: n kanssa.
on kaksi tärkeää adaptaatiota, joiden avulla C4-kasvit voivat tehdä tämän:
- ensinnäkin C4-kasvit käyttävät vaihtoehtoista entsyymiä hiilensidonnan ensimmäisessä vaiheessa. Entsyymi on nimeltään fosfoenolipyruvaatti (PEP) karboksylaasi, ja sillä ei ole oksigenaasiaktiivisuutta ja sillä on paljon suurempi affiniteetti CO2: een kuin rubiscolla. Nimensä mukaisesti” PEP-karboksylaasi ” entsyymi liittää hiilidioksidia yhdisteeseen nimeltä fosfoenolipyruvaatti (PEP).
- toiseksi, C4-kasveilla on erikoistunut lehtien anatomia, jossa on kaksi erilaista yhteyttävää solutyyppiä: mesofyllisolut (lehden ulkopinnalla lähellä ilmarakoa) ja nipukkasydämiset (lehden sisäpinnalla kaukana ilmarakosta). Rubisco sijaitsee niputupisoluissa, mutta ei mesofyllisoluissa.
näin tämä kaikki toimii:
PEP-karboksylaasi sijaitsee mesofyllisoluissa, lehtien ulkopinnalla lähellä ilmarakoa. Mesofyllisoluissa ei ole rubiscoa. Avantoon tuleva hiilidioksidi kiinnittyy nopeasti PEP-karboksylaasin vaikutuksesta 4-hiiliseksi yhdisteeksi eli malaatiksi kiinnittämällä hiilidioksidin PEP: hen. Tämän jälkeen malaatti kulkeutuu syvemmälle lehtikudokseen nipun tupisoluihin, jotka ovat molemmat kaukana avannosta (ja siten kaukana hapesta) ja sisältävät rubiscoa. Kun malaatti on bundle-vaippa-solujen sisällä, se dekarboksyloidaan vapauttaakseen pyruvaattia ja CO2: ta; RUBISCO kiinnittää sen Calvinin kiertoon, aivan kuten C3-kasveissa. Tämän jälkeen pyruvaatti palaa mesofyllisoluihin, joissa käytetään ATP: n fosfaattia elvyttämään PEP: tä. Niinpä C4-kasveissa C4-hiilensidonnan nettolisäyskustannukset ovat 1 ATP jokaista rubiscoon toimitettua hiilidioksidia kohti; C4-kasvit kuitenkin kuolevat kuivaan tilaan harvemmin kuin C3-kasvit kuivissa olosuhteissa.
alla oleva kuva havainnollistaa juuri kuvaamaamme reittiä:
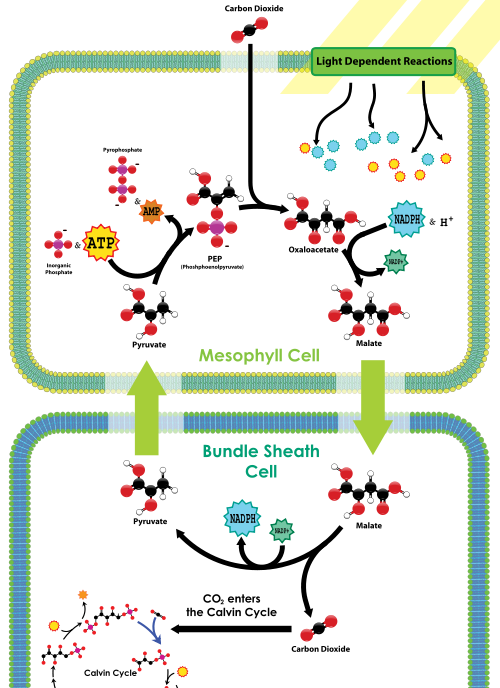
C4 hiilensidonta yleiskatsaus, Wikimediasta
Biol 1510 opiskelijan tulee muistaa C4: stä, että nämä kasvit ovat lisänneet CO2-konsentraatiomekanismin rubiscon ja Calvinin syklin ruokkimiseksi; mekanismi käyttää Pep-karboksylaasia aluksi 4-Hiilisen yhdisteen valmistamiseen, joka sitten vapauttaa CO2: ta rubiscoon lehtisoluissa, jotka altistuvat vähälle hapelle. Vaikka tämä mekanismi vähentää rubiscon oksigenaasiaktiivisuutta, sillä on ylimääräinen energiakustannus toisen ATP: n muodossa moolia kohden CO2 kiinteänä.
alla oleva (rajattu) video tarjoaa erinomaisen yleiskuvan C3: n ja C4: n fotosynteesin vertailusta, mutta ota huomioon seuraavat ongelmat tämän videon kanssa: video saa sen kuulostamaan siltä kuin RuBP katalysoisi omaa reaktiotaan CO2: n kanssa muodostaen 2: lle 3-hiili 3PG: n molekyylille sen sijaan, että se olisi yksi reaktion reaktioaineista. Tätä entsymaattista reaktiota katalysoi rubisco, ja juuri rubiscossa on oksignaasiaktiivisuutta, Ei RuBP! Videon perusteella kuulostaa myös siltä, että PEP katalysoi omaa reaktiotaan hiilidioksidin kanssa muodostaen 4-hiilimalaattia, kun todellisuudessa tätä reaktiota katalysoi PEP-karboksylaasi.
https://www.youtube.com/watch?v=HbLg4lMpUa
Jos haluat tietää enemmän, alla oleva video antaa perusteellisemman (vaikkakin hieman hitaamman) kuvan tästä prosessista:
C4 – ja C3-hiilensidonnan edut ja haitat
- C4-kasvit kasvavat paremmin kuin C3-kasvit kuumissa, kuivissa olosuhteissa, kun kasvien on suljettava ilmansa säästääkseen vettä-ilmaraot suljettuina, CO2-tasot lehtien sisäosissa putoavat ja O2-tasot nousevat.
- C3-kasvit kasvavat C4-kasveja paremmin viileissä, kosteissa olosuhteissa, kun kasvit voivat avata ilmansa, koska C3-kasvit eivät aiheuta C4-hiilen sitomisesta ATP: lle lisäkustannuksia.
fotosynteesi ja hengitys: peilikuvat
hapettuneen fotosynteesin ja aerobisen hengityksen kemialliset yhtälöt ovat täsmälleen toistensa vastakohtia.
fotosynteesin kokonaisnopeuden (alkutuotanto) ja kokonaishengitysnopeuden välinen tasapaino tarvitaan, jotta ilmakehän CO2-ja O2-pitoisuudet pysyvät vakaina.
eukaryooteilla sekä fotosynteesi että hengitys tapahtuvat organelleilla, joilla on kaksoiskalvot ja omat kehämäiset genominsa, jotka saavat alkunsa prokaryoottisina endosymbiontteina.
molemmissa prosesseissa on elektroninsiirtoketjut, kemiosmoosi ja ATP-syntaasi, jonka voimanlähteenä on protonimotiivinen voima.
Video screencasteissa käytetyt powerpoint-diat ovat Hiilensidontadiidijoukossa.