hoewel Rubisco verantwoordelijk is voor het overgrote deel van organische koolstof op het aardoppervlak, kan zijn oxygenase-activiteit de fotosynthetische efficiëntie aanzienlijk verminderen. Sommige planten hebben een manier ontwikkeld om de oxygenase activiteit van Rubisco te minimaliseren.
leerdoelstellingen
- Identificeer de omstandigheden die de oxygenase-activiteit van Rubisco verhogen
- beschrijf hoe de oxygenase-activiteit van Rubisco de fotosynthetische efficiëntie vermindert
- onderscheid C3 en C4 schema ‘ s voor koolstoffixatie
- wegen de voor-en nadelen van C3 versus C4
- vergelijk en contrast fotosynthese en ademhaling, en hun relatie in de Globale koolstof-en zuurstofcycli.
De oxygenase-activiteit van Rubisco belemmert de fotosynthetische efficiëntie
Rubisco heeft zowel oxygenase-activiteit als carboxylase-activiteit; het fixeert soms O2 aan RuBP in plaats van CO2. De oxygenase activiteit komt voor bij lage CO2, hoge O2 omstandigheden, en wordt uitgesproken bij hoge temperaturen. Als gevolg hiervan wordt organische koolstof geoxideerd, het tegenovergestelde van fotosynthese, die anorganische koolstof vermindert om organische koolstof te maken.
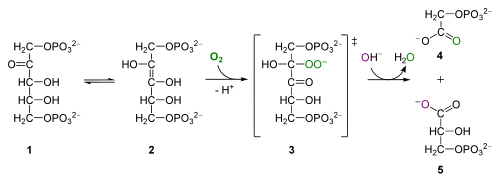
voor de nieuwsgierige oxygenase-activiteit van Rubisco, uit Wikipedia. RuBP (1) interconverteert naar een enolisomeer (2) dat zich combineert met zuurstof om het onstabiele tussenproduct te vormen (3) dat hydrolyseert in fosfoglycolaat (4) en 3PG (5)
de oxygenatie van RuBP produceert 2-fosfoglycolaat, een 2-koolstof toxische verbinding die een reeks reacties in het peroxisoom en mitochondriën ondergaat, waarbij CO2 vrijkomt en resulteert in verlies van organische koolstof en energieproductie. Dit proces heet fotorespiratie-een vreselijk misleidende naam voor studenten, omdat het niets met ademhaling te maken heeft en geen ATP oplevert. Alle Biol 1510 studenten moeten onthouden over fotorespiratie is dat het vermindert fotosynthetische efficiëntie, en dat het optreedt wanneer Rubisco oxygenates RuBP in plaats van carboxylating RuBP.
Rubisco evolueerde nog voor oxygenische fotosynthese, toen er geen zuurstof in de atmosfeer of in het oceaanwater was, dus was er geen selectie tegen oxygenase-activiteit. Niettemin, in meer dan 2 miljard jaar, heeft noch de natuur noch de menselijke genetische manipulatie de oxygenaseactiviteit van Rubisco kunnen elimineren of zelfs beduidend verminderen zonder ook de carboxylaseactiviteit te beïnvloeden.
C4 planten hebben een mechanisme ontwikkeld om CO2 te leveren aan Rubisco
om planten CO2 te laten opnemen, moeten ze structuren openen die stomata op hun bladeren worden genoemd, wat poriën zijn die gasuitwisseling mogelijk maken. Planten verliezen ook waterdamp door hun stomata, wat betekent dat ze kunnen sterven aan uitdroging in droge omstandigheden als ze hun stomata open te houden voor fotosynthese. Als reactie, planten sluiten hun stomata om uitdroging te voorkomen. De directe * gevolgen van gesloten stomata op korte termijn zijn verminderde CO2 concentratie (CO2 niveaus dalen als het wordt gebruikt voor fotosynthese) en verhoogde O2 concentratie (O2 niveaus stijgen als het wordt geproduceerd door fotosynthese). De stijgende O2 niveaus verhogen de snelheid van fotorespiratie (reactie van rubisco met zuurstof in plaats van koolstofdioxide), wanneer dan drastisch vermindert de efficiëntie van rubisco, die al een zeer langzaam-werkend enzym is. Dit betekent dat planten in droge omstandigheden risico lopen op uitdroging als ze hun stomata openen om gasuitwisseling te bevorderen, of het onvermogen om suiker te produceren als ze hun stomata gesloten houden om uitdroging te minimaliseren. (*het gevolg op lange termijn van permanent gesloten stomata is dood door verstikking als de mitochondriën geen zuurstof meer hebben om de ademhaling uit te voeren.)
veel planten die in warme en/of droge omstandigheden leven, hebben een alternatieve koolstoffixatieweg ontwikkeld om de efficiëntie van rubisco te verbeteren, zodat ze hun stomata niet zo veel open hoeven te houden, en zo verminderen ze het risico op sterfte door uitdroging. Deze planten worden C4-planten genoemd, omdat het eerste product van koolstoffixatie een 4-koolstofverbinding is (in plaats van een 3-koolstofverbinding zoals in C3 of “normale” planten). C4-planten gebruiken deze 4-koolstofverbinding om effectief CO2 rond rubisco te “concentreren”, zodat rubisco minder snel met O2 reageert.
Er zijn twee belangrijke aanpassingen waardoor C4-planten dit kunnen doen:
- Ten eerste gebruiken C4-planten een alternatief enzym voor de eerste stap van koolstoffixatie. Dit enzym wordt phosphoenolpyruvate (PEP) carboxylase genoemd, en het heeft geen oxygenaseactiviteit en heeft een veel hogere affiniteit voor CO2 dan rubisco. Aangezien de naam “Pep carboxylase” voorstelt, hecht het enzym CO2 aan een samenstelling genoemd phosphoenolpyruvate (PEP).
- ten tweede hebben C4-planten gespecialiseerde bladanatomie met twee verschillende soorten fotosynthetische cellen: mesofylcellen (aan de buitenkant van het blad, in de buurt van stomata) en bundel schede cellen (in het binnenste van het blad, ver weg van stomata). Rubisco bevindt zich in bundel schede cellen, maar niet in mesofyl cellen.
dit alles werkt:
pepcarboxylase bevindt zich in de mesofylcellen, aan de buitenkant van het blad bij de stomata. Er zit geen rubisco in de mesofylcellen. Het CO2 dat de stomata ingaat wordt snel door Pep carboxylase in een 4-koolstofsamenstelling, genoemd malaat gefixeerd, door het CO2 aan PEP vast te maken. Het malaat wordt dan dieper in het bladweefsel naar de bundel schede cellen getransporteerd, die zowel ver van de stomata (en dus ver van zuurstof) en rubisco bevatten. Eenmaal binnen de bundel schede cellen, wordt malaat gedecarboxylated om pyruvate en CO2 vrij te geven; de CO2 wordt dan bevestigd door rubisco als deel van de Calvijn cyclus, enkel zoals in C3 installaties. Pyruvate keert dan aan de mesophyll cellen terug, waar een fosfaat van ATP wordt gebruikt om PEP te regenereren. Dus in C4-installaties, C4 carbon fixation heeft een netto toegevoegde kosten van 1 ATP voor elke CO2 geleverd aan rubisco; C4-planten sterven echter minder vaak aan uitdroging dan C3-planten in droge omstandigheden.
de afbeelding hieronder illustreert de route die we zojuist beschreven hebben:
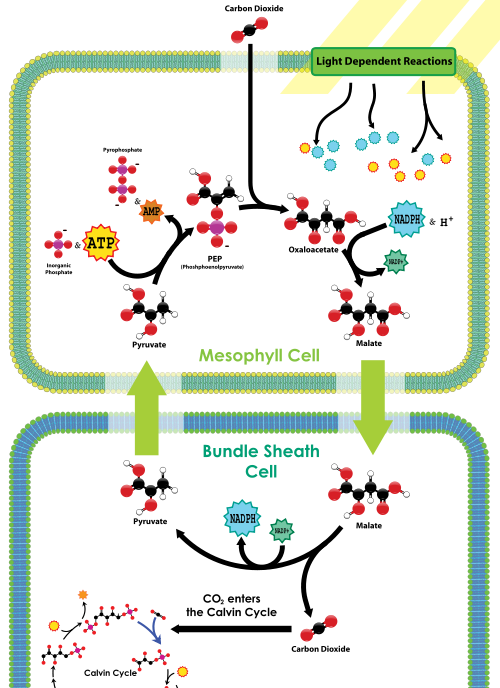
C4 carbon fixation overview, from Wikimedia
Wat Biol 1510 studenten moeten onthouden over C4 is dat deze planten een CO2 concentratie mechanisme hebben toegevoegd om rubisco en de Calvin cyclus te voeden; het mechanisme gebruikt PEP carboxylase om in eerste instantie maak een 4-koolstofverbinding, die dan CO2 afgeeft aan Rubisco in bladcellen die worden blootgesteld aan weinig zuurstof. Terwijl dit mechanisme de oxygenase-activiteit van rubisco vermindert, heeft het een extra energiekosten in de vorm van een ander vast ATP per mol CO2.
de (bijgesneden) video hieronder geeft een geweldige overzichtsvergelijking van C3 Versus C4 fotosynthese, maar wees je bewust van de volgende problemen met deze video: de video laat het klinken alsof RuBP zijn eigen reactie met CO2 katalyseert tot 2 moleculen van 3-koolstof 3PG, in plaats van een van de reactanten in deze reactie te zijn. Deze enzymatische reactie wordt gekatalyseerd door rubisco, en het is rubisco die oxygnase activiteit heeft, niet RuBP! De video laat het ook klinken als PEP katalyseert zijn eigen reactie met CO2 om 4-koolstofmalaat te vormen, terwijl in werkelijkheid deze reactie wordt gekatalyseerd door PEP carboxylase.
https://www.youtube.com/watch?v=HbLg4lMpUa
Als u meer wilt weten, geeft de video hieronder een grondigere (zij het wat langzame) illustratie van dit proces:
voor – en nadelen van C4-en C3-koolstoffixatie
- C4-planten groeien beter dan C3-planten onder warme, droge omstandigheden wanneer planten hun stomata moeten sluiten om water te besparen-met gesloten stomata vallen de CO2-niveaus in het binnenste van het blad en stijgen de O2-niveaus.
- C3-planten groeien beter dan C4-planten onder koele, vochtige omstandigheden wanneer planten hun stomata kunnen openen, omdat C3-planten niet de extra kosten van ATP van C4-koolstoffixatie hoeven te dragen.
fotosynthese en ademhaling: de chemische vergelijkingen voor Oxygene fotosynthese en aërobe ademhaling zijn precies het omgekeerde van elkaar.
een evenwicht tussen de Globale fotosynthesesnelheden (primaire productie) en de Globale ademhalingssnelheden is nodig om de atmosferische concentraties van CO2 en O2 stabiel te houden.
bij eukaryoten komen zowel fotosynthese als ademhaling voor in organellen met dubbele membranen en hun eigen cirkelvormige genomen, die zijn ontstaan als prokaryotische endosymbionten.
beide processen hebben elektronentransportketens, chemiosmose en ATP-synthase aangedreven door protonmotive force.
de PowerPoint-dia ‘ s die in de video screencasts worden gebruikt, bevinden zich in de Carbon fixation-dia-set.